
生物の体を構成する体細胞は、原則としてすべて同じ遺伝情報をもつ。それは、細胞がもつすべてのDNAが、細胞分裂前に一度複製されるからである。そうしないと、分裂ごとに遺伝情報を失っていくことになる。では、ここから遺伝情報を維持するためのメカニズムを説明していこう。
DNA複製の基本概念
DNA複製の基本的な概念は、ワトソンとクリックによるDNA二重らせん構造の提唱後すぐに示された(二重らせんを提唱した論文内で言及されている)。細胞分裂のときに遺伝情報が母細胞から娘細胞へ正確に受け継がれるためには,母細胞の全DNAが複製により倍加され、2コピーのDNAが2つの娘細胞に均等に分配される。DNAを正確に複製するためには,DNAの二重らせんを解いて、それぞれの鎖を鋳型として相補的なヌクレオチドを取り込めばよい。
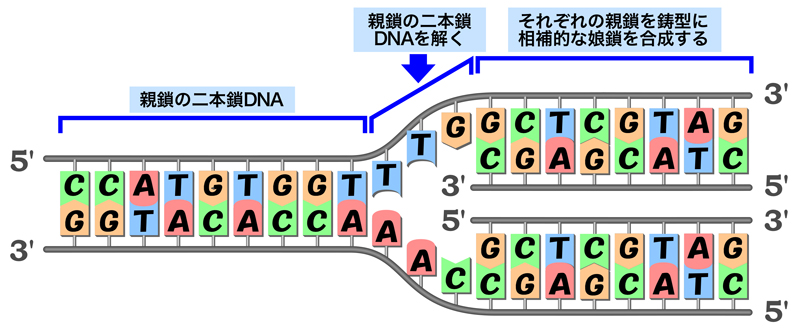
半保存的複製
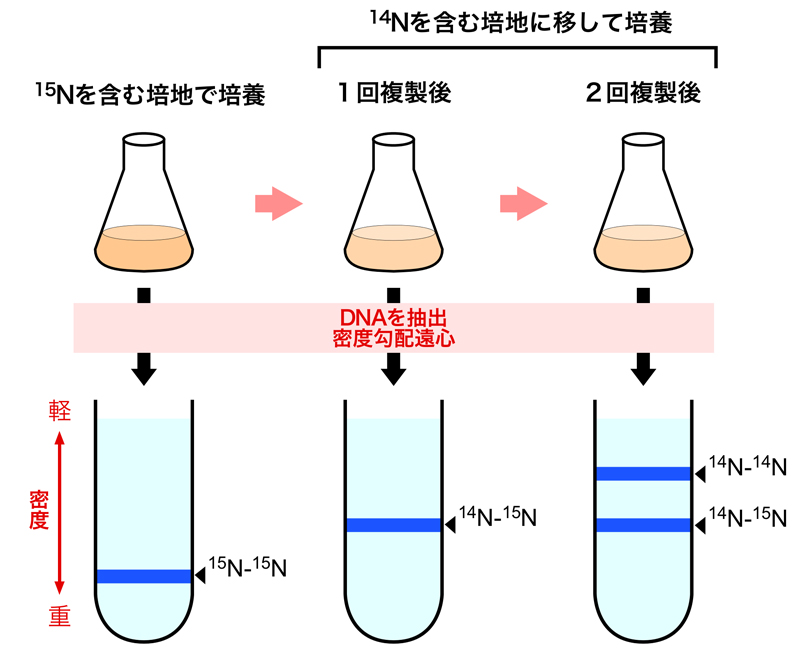
DNAの複製様式については3つのモデルが提唱されていた。「保存的複製モデル」では、親鎖の二本鎖DNAが解離して新生鎖合成の鋳型となったのち、2本の親鎖DNAは元の二重らせんに戻る。「半保存的複製モデル」では、複製後のDNAは鋳型となった親鎖と新生鎖のハイブリッドとして存在する。「分散的複製モデル」では、複製後のDNAの各々の鎖は、親鎖と新生鎖が混ざり合った状態となる。これらのうちどれが正しいかは、1958年のメセルソンとスタールの実験により明らかにされた。

彼らは、唯一の窒素源(塩化アンモニウムNH4Cl)として重い窒素の同位体である15Nを含む培地で大腸菌を培養し,DNAを15Nで標識した。この大腸菌を通常の軽い窒素14Nを含む培地に移して培養した後、様々なタイミングで大腸菌からDNAを抽出し、塩化セシウム(CsCl)の密度勾配遠心により解析した。このとき並行して大腸菌の増殖速度もチェックしておき、14Nを含む培地に移してから何世代増殖した(つまり何回分裂した)大腸菌からDNAを抽出したかも調べておいた。
DNA複製を1回行った大腸菌からのDNAを解析すると、大腸菌のDNAはすべて15N-14NのハイブリッドDNAであった。この結果により、「保存的複製モデル」は排除された。また、DNA複製を2回行った大腸菌からのDNAを解析すると、大腸菌からは14N-DNAと15N-14NハイブリッドDNAの両方が検出された。この結果により「分散的保存モデル」も排除された。このようにして、半保存的複製によりDNA複製が行われることが示されたのである。
DNA複製の概要
DNA複製は、複製起点とよばれるDNA上の特定の部位から開始される。DNA複製が開始するとき複製起点で二本鎖DNAは開かれ、そこからDNA合成が進行していく。このときDNA複製が行われて膨らんだように見える領域が、複製バブルとよばれる。そこからDNA複製が両方向あるいは一方向に進行、複製バブルは拡張していく。DNA合成が行われている複製バブルの末端部では、すでにDNA複製を終えた2本のDNAと未複製の親鎖DNAがY字型を成しており、この部分は複製フォークとよばれる。つまり、複製フォークが進行するにつれて複製バブルは拡張していく。そして、向き合う2つの複製フォークが出会ったところで複製バブルが融合し、その領域のDNA複製は終了する。

1つの複製起点からのDNA合成により複製される領域を、レプリコン(複製単位)という。原核生物のゲノムは環状DNAであり、1つの複製起点により複製される。つまり、1つの複製起点に由来する複製フォークが環状DNAの反対側で出会い、全ゲノムが複製される。してがって、原核生物のゲノムは1つのレプリコンということになる。一方、真核生物のゲノムは複数の染色体DNAで構成され、各染色体には多数の複製起点が存在する。したがって、真核生物のゲノムは多重レプリコンである。

DNAポリメラーゼ
DNA合成を行うのは、DNAポリメラーゼとよばれる酵素である。その作用の特徴は、以下の通りである。
- プライマーを必要とする。
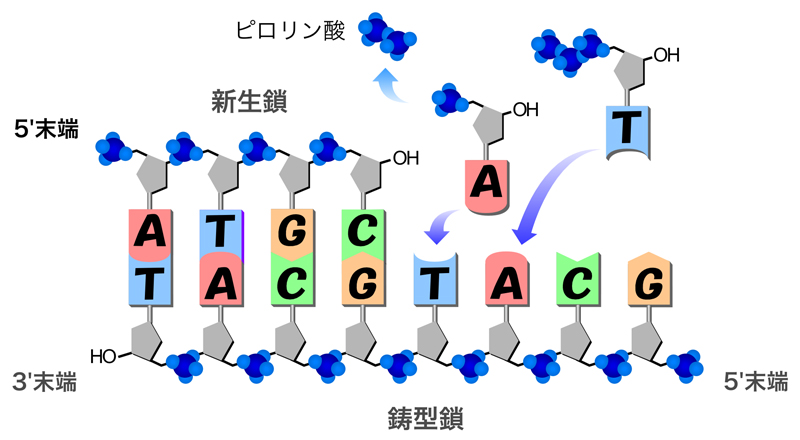
DNAポリメラーゼがDNA合成を開始するためには、プライマーとよばれる一本鎖の核酸(DNAまたはRNA)を必要とする。つまり、何もないところからDNAを合成することはできないのである。 - DNA合成の方向は、5’→3’方向である。
DNAポリメラーゼは、プライマーやDNA合成中の新生鎖の3’末端に鋳型DNAと相補的な塩基をもつヌクレオチドを取り込む酵素である。したがって、DNA合成は必ず5’から3’方向へと伸長していく。 - DNA合成の基質は、デオキシリボヌクレオシド三リン酸(dNTP)である。
DNA合成の基質は、デオキシリボヌクレオシド三リン酸(dNTP:dATP, dGTP, dCTP, dTTPの4種をまとめた総称)である。DNAポリメラーゼがDNA鎖の3’末端にヌクレオチドを付加する際、取り込む際には三リン酸のかたちで取り込む。そして、実際に付加する前にピロリン酸(二リン酸)を切り取り、残ったヌクレオシド一リン酸をDNAの3’-OH末端に付加する。 - dNTPの高エネルギーリン酸結合の加水分解により、必要なエネルギーを得る。
dNTPに含まれる3つのリン酸間の結合は、高エネルギーリン酸結合とよばれ、高いエネルギーをもっている。この加水分解において生じるエネルギーを利用して、DNAポリメラーゼは合成反応を実行している。
リーディング鎖とラギング鎖
DNAポリメラーゼがDNA合成を行う際、問題になるのはDNAを構成する2本の鎖が逆並行であるという点である。鋳型となるDNAの2本の鎖は逆平行なのに、DNAポリメラーゼは5’→3’方向にしかDNAを合成できない。したがって、それぞれの鋳型DNA鎖上での新生鎖の合成方向は逆になる。つまり、一方の鎖では複製フォークの進行方向とDNA合成の方向が同じになって問題ないのだが、もう一方の鎖では複製フォークの進行方向とDNA合成の方向が逆向きになってしまう。そこでDNA複製は、それぞれの鋳型DNA鎖で異なるDNA合成様式をとることにより、この問題を解決している。
下図の下側の鎖では、DNAの合成方向と複製フォークの進行方向は一致しており、DNA合成は連続的に行われる。このような合成様式のDNA鎖をリーディング鎖とよぶ。その反対側(下図の上側の鎖)では、DNAの合成方向と複製フォークの進行方向は逆になっている。そこで、複製フォークの進行方向と逆向きに岡崎フラグメントとよばれる短いDNA断片が合成され、これを複製フォークの進行方向に順次連結していく。このような合成様式のDNA鎖をラギング鎖とよぶ。岡崎フラグメントの長さは、原核生物では1,000〜2,000ヌクレオチド、真核生物では100〜400ヌクレオチドである。

