いよいよ転写の分子メカニズムです。
まずは、原核生物(バクテリア)の転写からみていきましょう。
原核生物のプロモーター
プロモーターというのは、RNAポリメラーゼが結合して転写の開始位置と方向を決定する重要な領域である。原核生物の様々なプロモーターの塩基配列を、転写開始点の位置を揃えて並べてみると、ある共通性が見えてくる。それは、転写開始点の上流になんとなく共通する領域があることである。

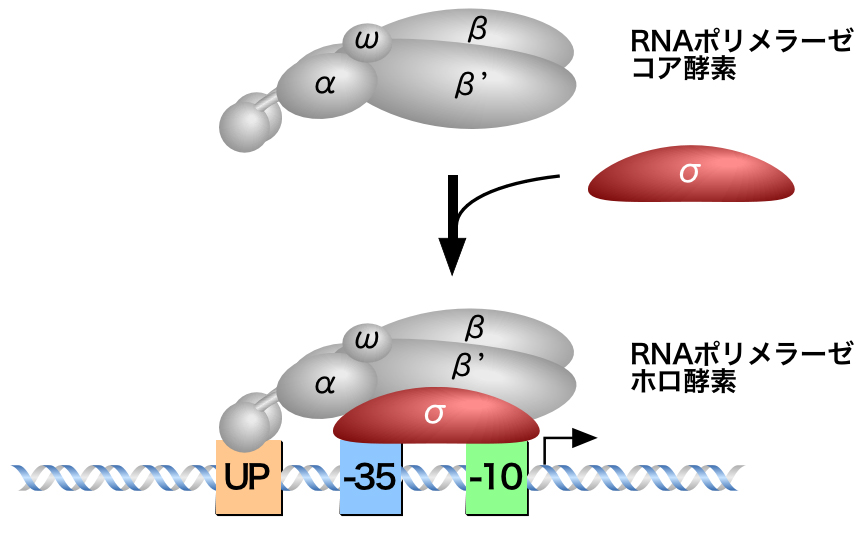
転写開始点(+1)のおよそ10 bp上流には、AT-richな塩基配列がみられる(上図の緑)。この領域は-10領域またはプリブナウボックスとよばれ、そのコンセンサス配列はTATAATである。また、およそ35 bp上流にも同様に保存された塩基配列があり(上図の水色)。この領域は-35領域とよばれ、そのコンセンサス配列はTTGACAである。さらに、-10領域と-35領域の間の長さは15〜19 bp(17 bpが最適)である。これらの塩基配列(特徴)が原核生物のプロモーターの保存されているのは、-10領域と-35領域を認識して結合するタンパク質が存在するからである。

また、rRNA遺伝子のような強力なプロモーターでは、転写開始点(+1)のおよそ40〜60 bp上流にUP配列とよばれるAT-richな塩基配列が存在する。
原核生物のRNAポリメラーゼ
原核生物では、1種類のRNAポリメラーゼがすべての遺伝子の転写を行っている。原核生物のRNAポリメラーゼは、複数ポリペプチド(サブユニット)で構成されており、それぞれが異なる役割を担う。2つのαサブユニットが酵素の集合・プロモーターの認識・転写因子との結合を、βサブユニットとβ’サブユニットが活性中心としてRNAの合成を、ωサブユニットが酵素の集合と安定化を担っている。そして、σサブユニットがプロモーターへの特異的結合を担う。
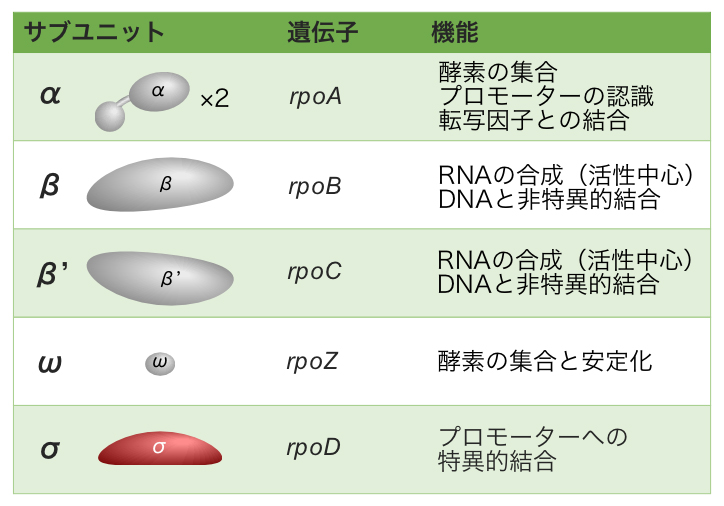
これらのサブユニットが集合してRNAポリメラーゼという酵素を形づくられる。2つのαサブユニットとβ・β’・ωサブユニットが1つずつ集まって、RNAポリメラーゼのコア酵素が形成される。そして、このコア酵素にσサブユニットが結合して、RNAポリメラーゼ ホロ酵素となる。
コア酵素はσサブユニットをもっていないが、RNAの合成を効率よく行うことができる。したがって、鋳型DNAと混ぜてin vitroで転写反応を行うと、RNAポリメラーゼコア酵素はDNA上のどこからでも転写を開始でき、ランダムな長さの転写産物RNAを生じる。
![]()
一方、ホロ酵素はσサブユニットをもっているため、プロモーターを特異的に認識できる。したがって、プロモーター上の正しい部位から転写を開始でき、in vitro転写では一定の長さの転写産物RNAを生じる。
![]()
このように、σサブユニットはRNAポリメラーゼコア酵素をプロモーターへと導く重要な役割を担う。それは、σサブユニットがプロモーターを特異的に認識して結合できるからである。σサブユニットのドメイン2.4が-10領域を、ドメイン4.2が-35領域を特異的に認識する。σサブユニットの構造とプロモーターの塩基配列がピッタリ合うと、σサブユニットによるプロモーターの結合特異性は上昇し、効率的にプロモーターが認識されることになる。だから、-10領域と-35領域の塩基配列や、その間の長さが保存されているのである。また、上流のUP配列はαサブユニットのC末端ドメインが認識する。これにより、プロモーターが効率的に認識され、転写活性も上昇するのである。