
現在の遺伝子組換え技術には欠かせないツールとなっているのが、制限酵素。この頁では、制限酵素についてもう少しだけ深く見てみましょう。
制限-修飾系
制限酵素は、制限-修飾系(restriction–modification system)の研究で発見された。
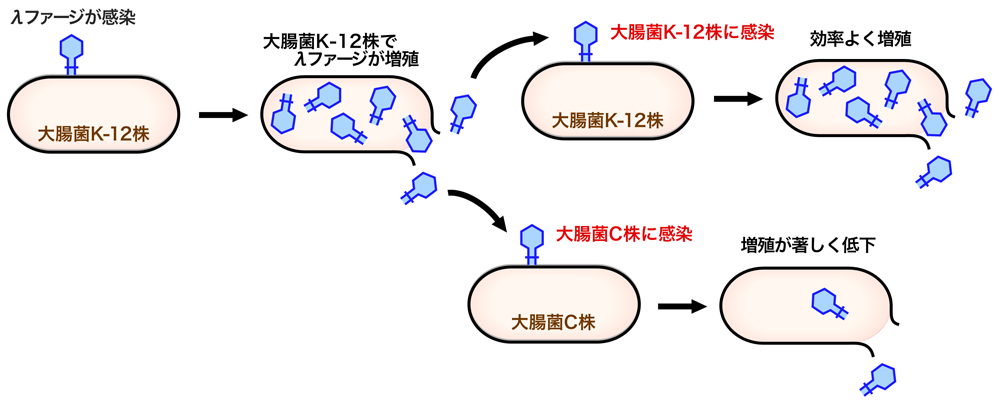
制限-修飾系とは、ある型のバクテリアに感染して増殖したバクテリオファージを、別の型のバクテリアに感染させたときに起こる現象である。例えば、大腸菌K-12株に感染して増殖したλファージを同じ大腸菌K-12株に感染させるとλファージはよく増えるが、同じλファージを大腸菌C株に感染させると、新しい宿主により増殖が制限されてλファージの増殖は大きく低下する。しかし、大腸菌C株で増殖できたλファージをさらに大腸菌C株に感染させると、増殖能は元に戻る。
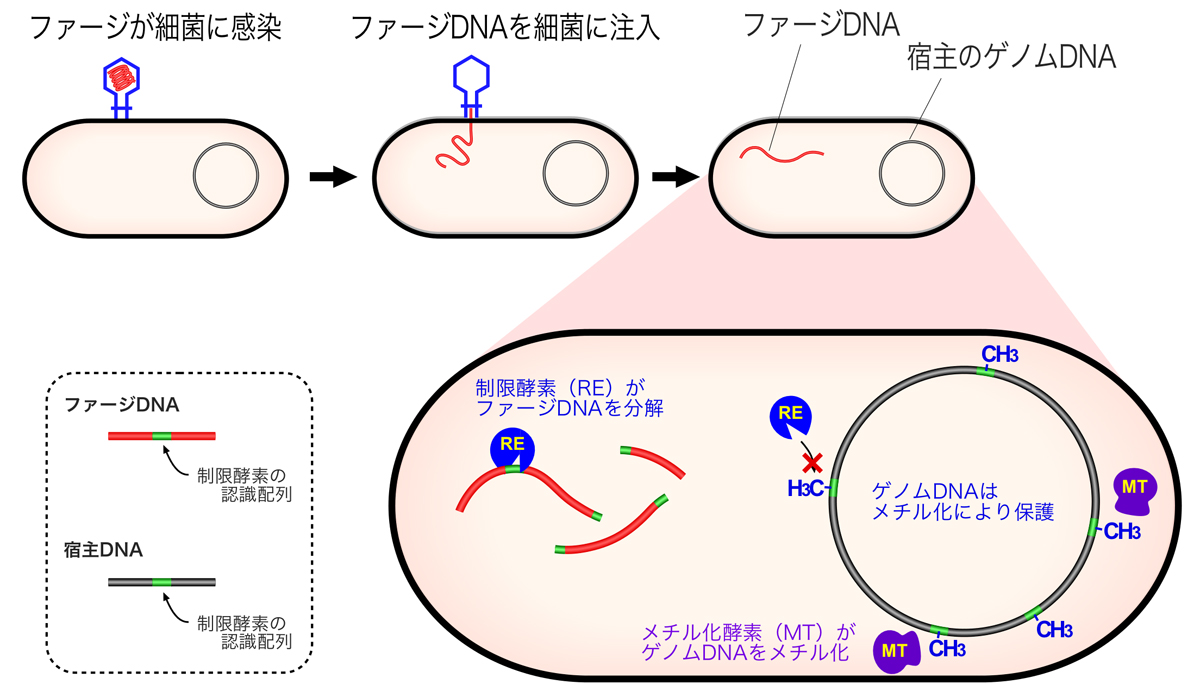
各種・各型のバクテリアは、特定の塩基配列を認識して切断する制限酵素をもっている。バクテリアは、外来のDNAと区別して自身のゲノムDNAを保護するために、制限酵素と同じ配列を認識してメチル化するメチル化酵素(メチルトランスフェラーゼ)をもっている。このメチル化が”修飾”であり、制限酵素の認識配列をメチル化することによって、自身のゲノムDNAを制限酵素による制限(制限酵素による分解)から回避させるのである。
制限酵素の命名
制限酵素の名称には、由来するバクテリアの属名・種名・株(血清型)・発見の順序に基づいた決まりがある。まず、属名の最初の1文字と種名の最初の2文字を並べる。例えば大腸菌なら学名がEscherichia coliなので、大腸菌に由来する制限酵素にはEcoが使われる。学名には通常斜体が使われるのでEcoの部分には斜体を使う慣わしがあったが、2003年に制限酵素の表記法についての取り決めが発表され1)、最近は斜体にしないようである。続いて、もしあれば株(血清型)を示し、最後に発見の順序をローマ数字で表す。よって、大腸菌Escherichia coliのRY13株に由来する最初に発見された制限酵素は、EcoRIと命名された。
同様に、
- インフルエンザ菌Haemophilus influenzaeの血清型dに由来する3番目に発見された制限酵素は、HindIII
- セラチア菌Serratia marcesensに由来する1番目に発見された制限酵素は、SmaI
- クレブシエラ肺炎桿菌Klebsiella pneumoniaeに由来する1番目に発見された制限酵素は、KpnI
となる。
制限酵素の認識配列
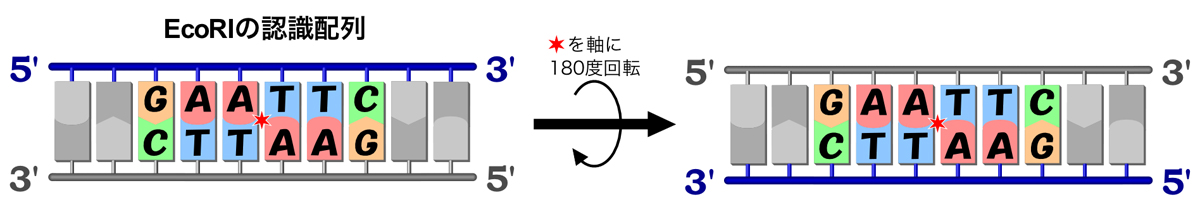
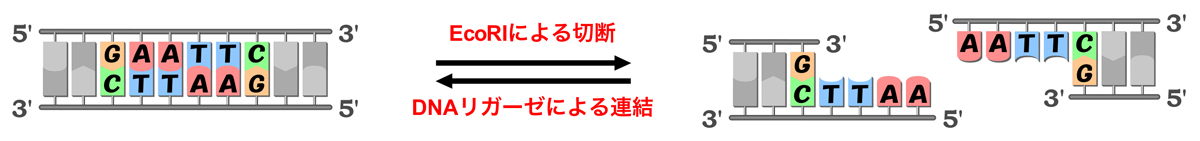
DNAの2本の鎖は逆並行であるため、DNAの構造は2回転対称となる。つまり、180度ひっくり返しても同じ構造なのだ。そして制限酵素の認識部位も同様で、180度ひっくり返しても同じ塩基配列となるような対称性をもつ。例えば、EcoRIは5’-GAATTC-3’という塩基配列を認識して切断する。この配列は、下図のように180度ひっくり返しても同じ配列/構造となる。認識配列の最初の3塩基と続く3塩基が相補的な関係になっているので、こうなるのだ。このような配列を、パリンドロームという。
認識配列の長さも酵素によって異なり、4塩基、5塩基、6塩基、8塩基を認識するものがある。認識する配列の長さによってDNA中の出現頻度が異なり、4塩基認識の場合は44=256塩基対に1回、6塩基認識の場合は46=4096塩基対に1回となる。
制限酵素による切断
制限酵素による切断では、はじめに制限酵素がDNAの糖-リン酸骨格に非特異的に接触し、その後DNAに沿って移動しながら配列をスキャンする。EcoRV場合、1回の結合で2×106塩基対を1秒あたり1.7×106塩基対の速度でスキャンすると言われる。特定の塩基配列を見つけると、制限酵素は認識配列中の両方の鎖に切れ目を入れ、DNAの二本鎖の切断が起こる。切断の際は、ホスホジエステル結合の部分で加水分解(水分子が付加されて切断)が起こり、2つのヌクレオチド間に切れ目が入る。結果、リン酸基をもつ5’末端と水酸基をもつ3’末端を生じる。
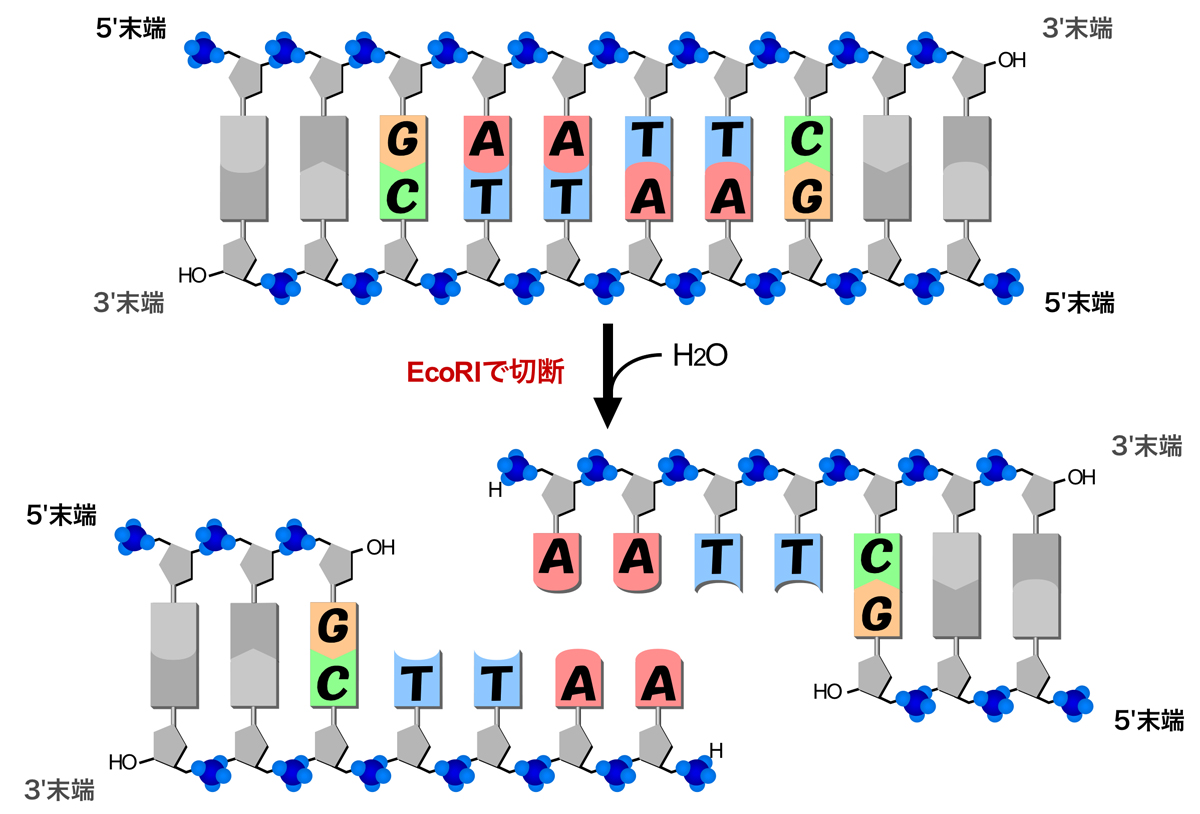
制限酵素による切断では、切れ目の入る位置によって突出末端を生じる。しかも認識配列がパリンドロームであるため、生じた2つの突出末端は相補的な塩基配列となり、再び連結することができる。連結の際は、DNAリガーゼという酵素が脱水縮合(水分子を取り除いて連結)を行う。例えばEcoRIによる切断では、5’-AATT-3’という一本鎖の塩基配列がDNAの5’末端に突出する。EcoRIによる切断部位にはすべて同じ塩基配列/構造の末端を生じるため、EcoRIによる切断部位はすべてDNAリガーゼによって連結可能となる。そのDNAが由来する生物種に関係なく連結可能であり、こうした事実に基づいて遺伝子組換え技術が確立された。
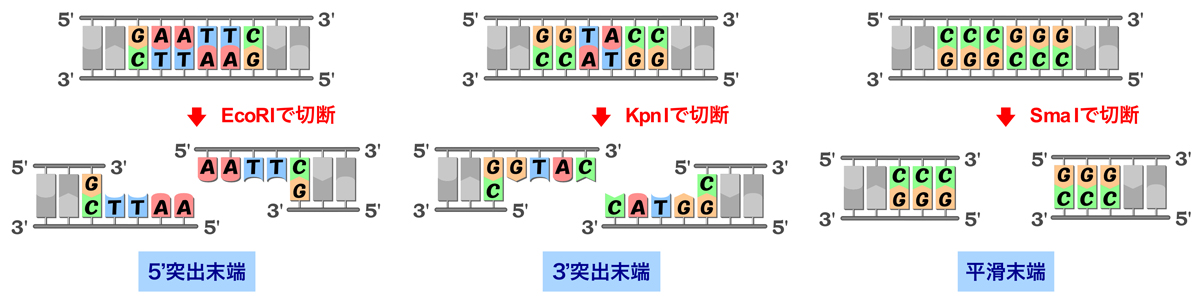
切断末端の形状や塩基配列は制限酵素によって異なる。HindIIIはEcoRIと同様に5’末端が突出した末端を生じるが、KpnIやPstIのように3’末端が突出したり、EcoRVやSmaIのように平滑末端を生じる制限酵素もある。平滑末端を生じる場合は末端間に相補性がないため、異なる制限酵素による切断末端間での連結も可能になるので、注意しよう。
メチル化による修飾
制限-修飾系の修飾を担うのは、DNAのメチル化である。バクテリアは制限酵素に対応するメチル化酵素をもっており、制限酵素EcoRIは5’-GAATTC-3’をに意識して切断し、対応するEcoRIメチラーゼは同じ配列を認識してメチル化し5’-GAm6ATTC-3’を生じる。同様に、制限酵素HindIIIは5’-AAGCTT-3’をに意識して切断し、対応するHindIIIメチラーゼは同じ配列を認識してメチル化し5’-m6AAGCTT-3’を生じる。
メチル化酵素は、制限酵素と一体となっている場合もあるし、分離している場合もある。共通するのは、メチル化のためのメチル基供与体としてS-アデノシルメチオニン(SAM)を必要とすることである。
アイソシゾマーとネオシゾマー
複数の制限酵素が同じ塩基配列を認識することがある。このような制限酵素をアイソシゾマー(isoschizomer)という。また、同じ配列を認識するけれども切断末端の形状が異なるものをネオシゾマー(neoschizomer)という。アイソシゾマーの中でも、最初に発見された制限酵素をプロトタイプ(prototype)という。
例えば、BclIとFbaI(どちらも5’-TGATCA-3’を認識)はアイソシゾマーである。また、NaeIとNgoMIV(どちらも5’-GCCGGC-3’を認識)もアイソシゾマーであるが、NaeIは5’-GCC/GGC-3’、NgoMIVは5’-G/CCGGC-3’(それぞれ/の部分に切れ目が入る)にように切断するため、これらはネオシゾマーでもある。
さらに、認識配列の中央部(対称軸辺り)が一致する制限酵素もあり、切断末端の配列/形状が一致する場合がある。例えばBamHI(5’-GGATCC-3’を認識)とBglII(5’-AGATCT-3’を認識)は、どちらも認識配列中央の5’-GATC-3’が一致しており、どちらも5’-GATC-3’が5’突出末端となる。また、Sau3AIは認識配列が5’-GATC-3’の4塩基であり、切断によって5’-GATC-3’の5’突出末端を生じる。したがって、これらの制限酵素による切断末端は互いに連結可能ということになる。BamHIとBglIIによる切断末端を連結した場合、連結後の塩基配列はBamHIやBglIIでは認識されないことになるが、Sau3AIでは認識できる。

制限酵素の分類
制限酵素(制限-修飾系)は、分子構造・認識配列・切断の様式、活性に必要な補助因子によって4つのクラスに分類される。制限(R: restriction)、修飾(M: modification)、認識(S: specificity)の3つドメインが、3つのポリペプチドに別々に存在したり、2つあるいは1つのポリペプチドに共存したりする。ほぼ全ての制限-修飾系に共通するのは、そのヌクレアーゼ活性にMg2+のような二価の金属イオンを必要とすること、メチル化のためのメチル基供与体としてSAMを必要とする点である。
- タイプI(例:EcoKIやEcoBI)
3つのhsd (host specificity determinant)遺伝子によってコードされるタンパク質で構成される。通常、2つのhsdR(制限サブユニット)、2つのhsdM(修飾/メチル化サブユニット)、1つのhsdS(配列認識サブユニット)の5量体であり、活性にはMg2+、SAM、ATPが必要である。タイプIの制限酵素は、離れた2つの配列を認識し(EcoKIの認識配列はAAC(N6)GTGC)、認識部位から遠く離れた部位(距離は可変的)を切断する。 - タイプII(例:EcoRIやHindIII)
組換えDNAのツールとしてもよく使われる、制限酵素の中でも最大のグループである。一般にホモ二量体で作用し、ヌクレアーゼ活性にはMg2+のみが必要であり、認識配列内または近くにある決まった位置で切断する。タイプIIの制限酵素にはさらに、その特徴によって以下のようなサブタイプに分類されるが、各制限酵素がどれか1つだけに分類されるとは限らない。・タイプIIP:認識とヌクレアーゼのドメインが一体化しており、パリンドローム配列を認識して、認識配列内部を切断する(例:EcoRIやHindIII)・タイプIIS:認識とヌクレアーゼのドメインが分離しており、非対称な塩基配列を認識して、片側の少し離れた部位を切断する(例:HphIやFokI)・タイプIIA:非対称な配列を認識する(例:FokI)
・タイプIIB:認識配列の両側を切断する(例:BcgI) などなど… - タイプIII(例:EcoP1IやEcoP15I)
非対称な配列を認識し、逆方向の(向き合った)2つの認識配列が必要。
一方の認識配列の25~27塩基対下流の部位を切断する。
ヌクレアーゼ活性には、Mg2+とATPが必要。 - タイプIV
メチル化DNAのみを切断するが、配列の特異性は低いらしい。
参考文献
1) Roberts R.J. et al. (2003) Nucleic Acids Res. 31, 1805-1812
