
真核生物のRNAポリメラーゼIIは、タンパク質のアミノ酸配列をコードするmRNAの転写を担う。ここでは、RNAポリメラーゼIIによって転写される遺伝子のプロモーターの構造について解説しよう。
4つのコンセンサス配列
RNAポリメラーゼIIによって転写される遺伝子のプロモーターといえば、まずはじめにTATAボックスを連想するだろう。ところが、TATAボックスをもつプロモーターは、全プロモーターの10~15%あるいはそれ以下ともいわれる。しかし、プロモーターに含まれる配列はこれだけではない。
プロモーターといっても、正しい位置からの適切な転写開始に必要なコアプロモーターと、そのすぐ上流に存在してその本質的な活性を高める領域とがある。後者に含まれるのが、CAATボックスやGCボックスである。このページでは、コアプロモーターの方に焦点をあてる。
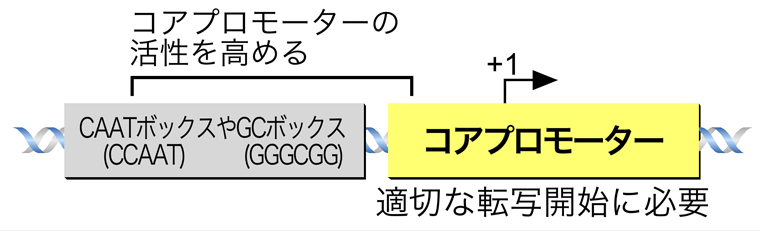
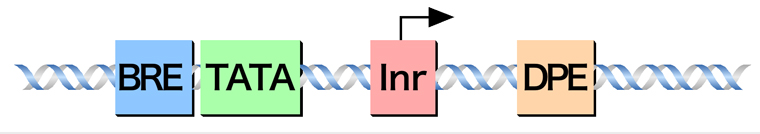
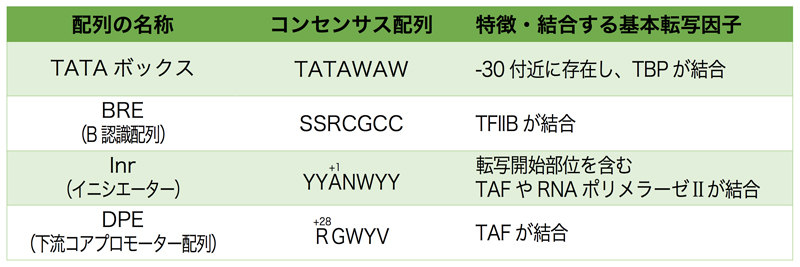
コアプロモーターに含まれる主要なコンセンサス配列は、以下の4つである。転写開始点より上流25〜35塩基対の位置にあるTATAボックス、基本転写因子TFIIBが結合部位であるB認識配列(BRE)、転写開始点を含む位置にあるイニシエーター(Inr)、転写開始点より下流にある下流コアプロモーター配列(DPE)である。どれも基本転写因子が結合する配列であるが、その詳細は次ページの基本転写因子の項で説明する。
その他のコンセンサス配列
最近では、上記4つのコンセンサス配列以外にも、コンセンサス配列が同定されているので紹介しよう。まずは、2002年のDrosophilaプロモーターの解析で見つかった10番のモチーフであるMTE (motif ten element)である。そのコンセンサス配列は、CSARCSSAACである。ちなみに、同解析においてTATAボックスは3番、Inrは4番、DPEは9番のモチーフとして同定されている。
さらに、DCE(downstream core element)は、DCEI(+6~+11:CTTC)、DCEII(+16~+21:CTGT)、DCEIII(+30〜34:AGC)の3つのサブ配列から構成される。ちなみにMTEもDCEも、どちらも基本転写因子TFIIDによって認識される。

また、BREについては、もともとはTATAボックスの上流に見出されていたが、その後TFIIBはTATAボックスの上流と下流の両方に結合することが判明し、従来のBREをBREUとし、下流側をBREd(コンセンサス配列はRTDKKKK)としている。
とても大事なポイント
RNAポリメラーゼIIにより転写されるプロモーターのコンセンサス配列を挙げてきたが、転写開始にはこれら全てが必要なのだろうか?冒頭にも書いたが、TATAボックスを含むのは全プロモーターの10〜15%あるいはそれ以下である。つまり、これらのコンセンサス配列はどれもプロモーター機能にとって必須ではないが、ほとんどのプロモーターはこれらの配列のうち2つないし3つだけをもつことにより、プロモーターとして機能するのである。
これらの配列の組み合わせについて、少しまとめる。TATAボックスは、単独でも転写を開始できるが、Inrなどの他の配列とも組み合わせても機能する。DCEは高頻度にTATAボックスと組み合わされるようである。Inrはもっとも頻繁に使われる配列であるが、単独では作用が弱く、通常は他の配列と組み合わせて存在する。MTEとDPEは、どちらも単独では機能しないが、Inrと組み合わさることにより活性をもつ。また、ほとんどのDPEやMTEを含むプロモーターは、TATAボックスを持たない。こんな感じで、組み合わせが重要なのである。
