ポリペプチド鎖の伸長は原核生物と真核生物とでよく保存されたメカニズムで行われている。しかし、翻訳開始のメカニズムは、原核生物と真核生物で大きく異なる点がある。そこで、翻訳開始のメカニズムを紹介しよう。ついでに、翻訳の終結についても、最後に簡単に解説したい。
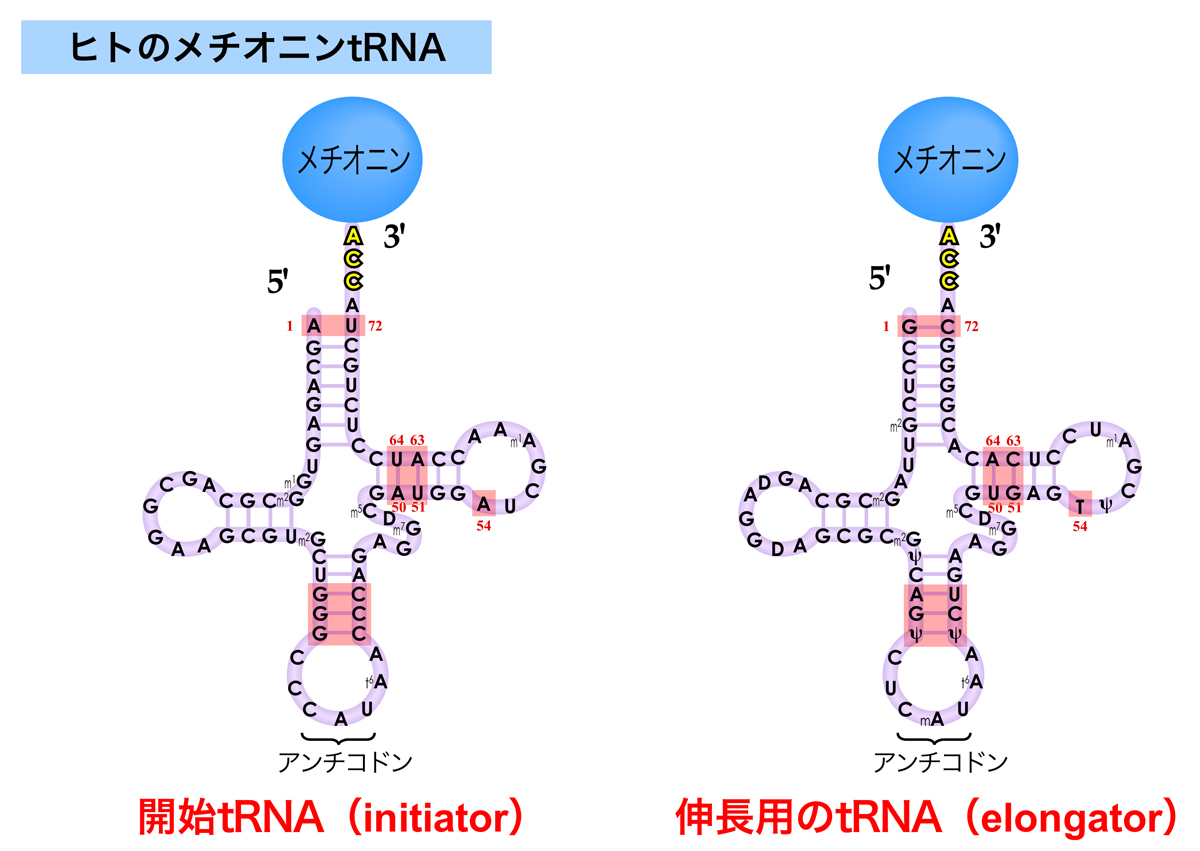
2つのメチオニンtRNA
タンパク質合成(翻訳)は、メチオニンをコードするAUGコドンで開始する。そこで、生物は開始用のメチオニンを運ぶtRNA(開始tRNA:initiator tRNA)と伸長用のtRNA(伸長tRNA:elongator tRNA)の2種類をもっている。開始tRNAは、リボソーム小サブユニットのP部位に直接結合し、翻訳開始に使うメチオニンだけを運ぶ。一方、伸長tRNAはポリペプチド鎖の伸長中に取り込まれるメチオニンを運び、リボソームのA部位に結合したのちにP部位へと移動する。その違いは、それぞれのtRNAが開始因子と伸長因子のどちらと結合するかによる。
また、細菌やミトコンドリアや葉緑体の開始tRNAに結合したメチオニンは、その後ホルミル化されてN-ホルミルメチオニンとなる。つまり、細菌の開始tRNAはN-ホルミルメチオニンを運ぶ。しかしほとんどの場合、ホルミル基とメチオニン残基は翻訳後に除去される。
細菌の開始tRNAには、伸長tRNAにはない以下の特徴がある。
- 受容ステムにある1Cと72Aの塩基間に水素結合がない。
ここを認識してホルミル化酵素(methionyl tRNA transformylase)がメチオニンをホルミル化する。伸長tRNAには1G:72Cの塩基対がある。 - アンチコドンループのステム部分に3連続のG:C塩基対がある。
- Dループのステム部分にプリン11とピリミジン24の塩基対がある。
伸長tRNAではピリミジン11とプリン24 - 50:64と51:63塩基対も伸長tRNAとして機能させないために重要。

真核生物の開始tRNAには、伸長tRNAにはない以下の特徴がある。
- 受容ステムにA1:U72塩基対がある。
開始tRNAのA1:U72塩基対は古細菌から真核生物まで不変で、唯一の例外は分裂酵母S. pombeのψ1:A72。伸長tRNAでは、この位置はすべての生物でG1:C72。 - アンチコドンループのステム部分に3連続のG:C塩基対がある。
- TψCループに、その名の由来となっている54番目のTがない。
伸長tRNAにおけるこの54番目のTは、伸長因子eEF1Aによる認識に重要 - A50:U64とU51:A63塩基対は、伸長因子eEF1Aの結合から守ることで、伸長tRNAとして機能させない。また、植物と菌類では、64番目の残基に2′-O-ribosylphosphateという修飾を受け、これが伸長tRNAとして機能させないために重要らしい。
原核生物の翻訳開始
原核生物のさまざまなmRNAの塩基配列を開始コドンの位置を揃えて並べると、開始コドンの約10ヌクレオチド上流にグアニン(G)やアデニン(A)に富む配列があることがわかる。これらの配列は、リボソーム小サブユニットを構成する16S rRNAの3’末端部分と一部相補的になっている。これを見出した研究者の名をとって、この配列はシャイン・ダルガーノ配列(Shine-Dalgarno配列)(AGGAGGU)とよばれる。シャイン・ダルガーノ配列は、16S rRNAの3’末端と相補的に結合することによりリボソーム結合部位として機能し、AUG開始コドンの位置を特定するのに役立っている。

原核生物の翻訳開始では、N-ホルミルメチオニンを運ぶ開始tRNAがリボソーム小サブユニットのP部位に結合して予め複合体を形成している。この複合体中の16S rRNAがシャイン・ダルガーノ配列と塩基対を作って結合すると、リボソームがちょうど開始コドンの部分に配置され、開始コドンから適切に翻訳が始まる。
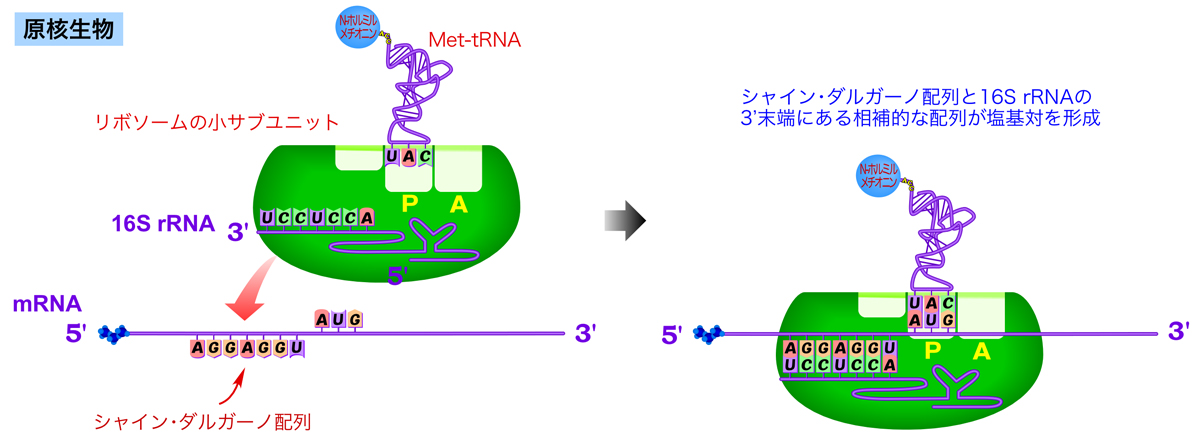
このように、シャイン・ダルガーノ配列があればそこにリボソームが結合して翻訳を始められるため、原核生物は1つのmRNA中に複数のポリペプチドをコードすることができるのである。
真核生物の翻訳開始
真核生物には、原核生物のシャイン・ダルガーノ配列のような明確なリボソーム結合配列はない。そこで、真核生物のmRNAに特徴的な5’キャップ構造を利用して、リボソームを呼び寄せる。
原核生物同様、メチオニンを運ぶ開始tRNAがリボソーム小サブユニットのP部位に結合して複合体を形成し、これがmRNAの5’キャップに結合する。mRNAの結合したリボソームは、mRNAに沿って5’→3’方向に移動し、AUG配列を探索する。mRNAの90%では、最初に現れるAUG配列を開始コドンとして翻訳が始まる。また、AUG周辺の塩基配列が翻訳開始を促進するのに重要である。一部のmRNAには、開始コドン周辺にコザック配列(Kozak配列)(GCCRCCAUGG)とよばれるコンセンサス配列が存在し、これがあると翻訳の効率が上昇する。逆に、コンセンサス配列と大きく違うと、リボソームが通過して次に現れるAUG配列を開始コドンとするらしい。
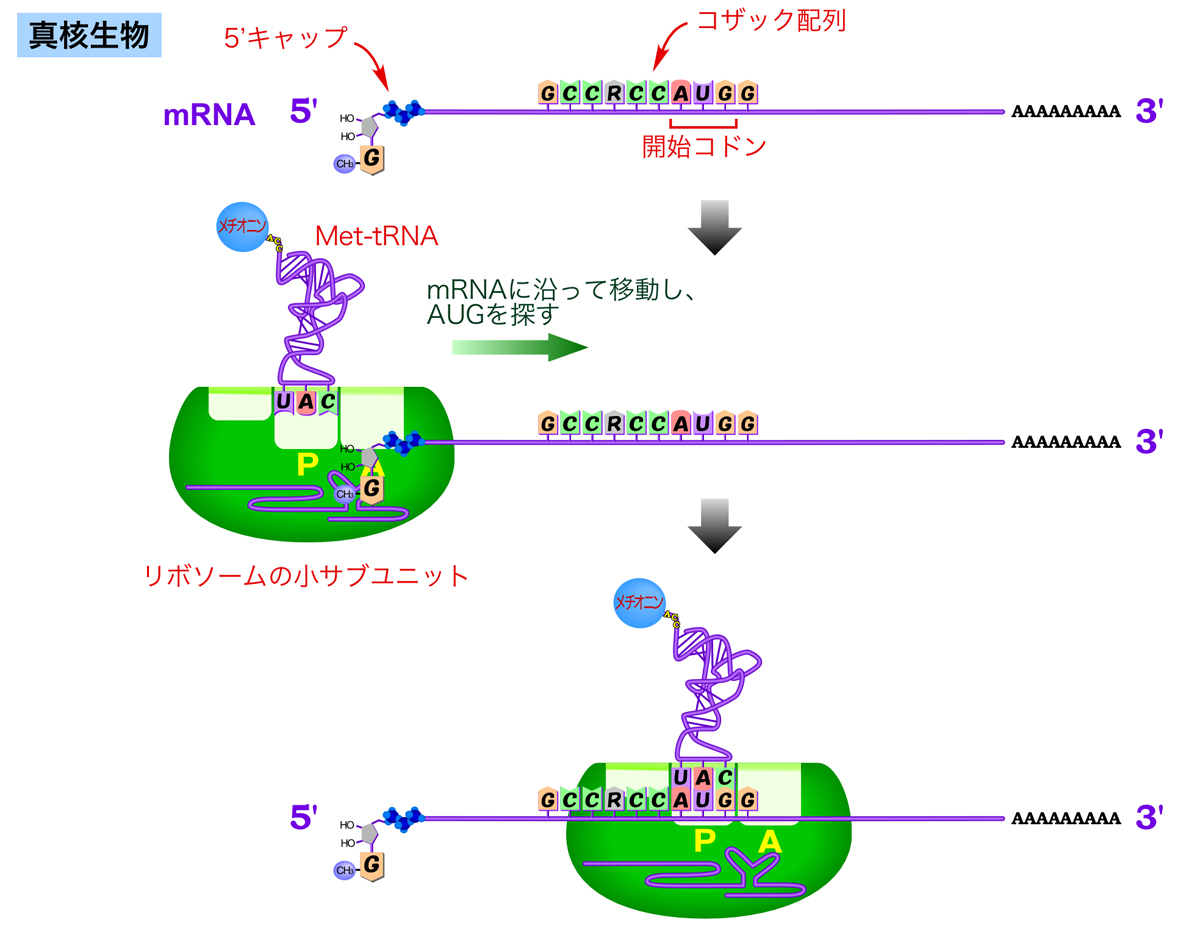
このように、mRNAの5’キャップ構造からスキャンして開始コドンを見つけるため、真核生物では1つのmRNA中にコードできるポリペプチドは原則として1つだけである(5’キャップに依存しない翻訳開始のしくみももっている)。
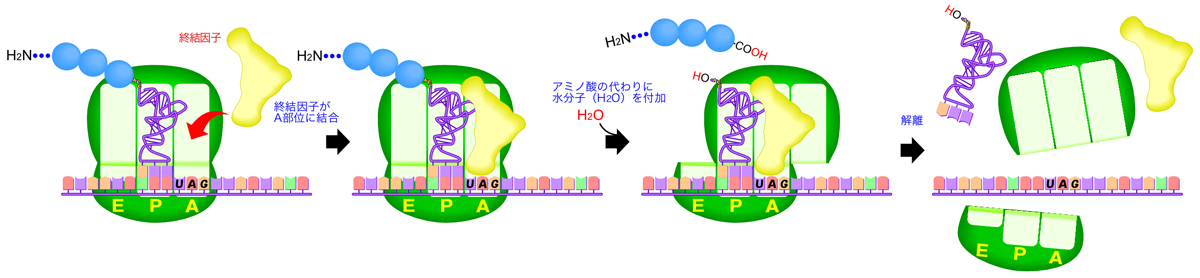
翻訳の終結
リボソーム上でポリペプチド鎖を伸長していく過程で、A部位に終止コドン(UAA, UAG, UGA)のいずれかが現れると、翻訳は終結する。ここで登場するのが、終結因子(release factor)である。終結因子は全体の立体構造がtRNAと類似しており、A部位に終始コドンが来ると、アミノアシルtRNAに代わってA部位に結合する。
原核生物には、RF1とRF2という2つのクラスI終結因子があり、RF1はUAGとUAA、RF2はUGAとUAAを識別する。クラスI終結因子には、終止コドンを識別する3アミノ酸の配列があり、ペプチドアンチコドンとよばれる。一方真核生物のクラスI終結因子にはeRF1しかなく、この一種類ですべての終止コドンに対応する。
終結因子がA部位に結合すると、ポリペプチド鎖のC末端にはアミノ酸の代わりに水分子を付加され、ポリペプチド鎖のC末端にカルボキシル基(COOH基)が形成されてtRNAから解離する。最後に、クラスII終結因子(原核生物のRF3、真核生物のeRF3)が、リボソームからクラスI終結因子が解離するのを促進する。