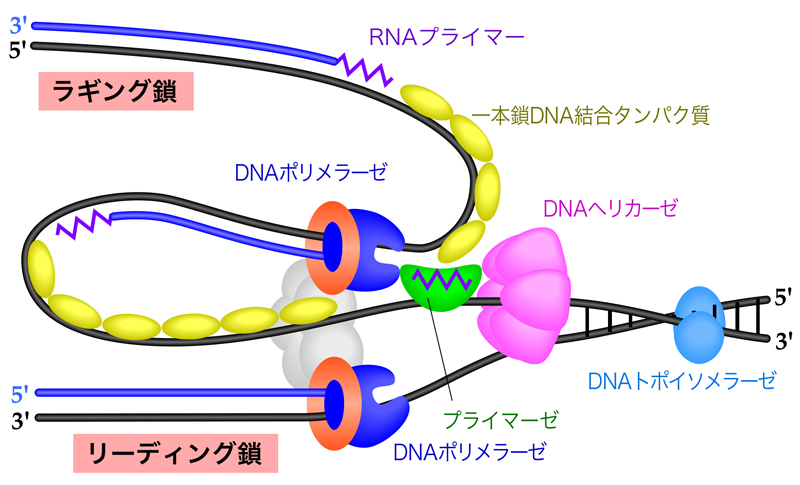
複製フォークでは、さまざまな因子が役割分担して協調的にはたらき、DNA複製を行っている。しかも、リーディング鎖とラギング鎖ではDNA合成様式が異なるにもかかわらず、複製フォークでは効率よくDNA合成が進行し、長大なゲノムDNAがすべて複製されるのである。このページでは、複製フォークで行なわれているDNA複製の分子機構を説明していこう。
DNAポリメラーゼ
DNA合成を行うDNAポリメラーゼにもいろいろあるが、下の表に生物がもつ主要なDNAポリメラーゼをリストアップした。この中でDNA複製に関与するのは、原核生物ではDNAポリメラーゼIとIII、真核生物ではDNAポリメラーゼα, δ, εである。
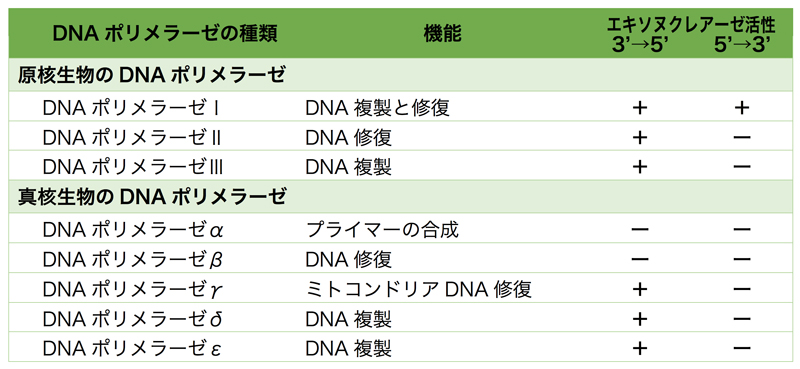
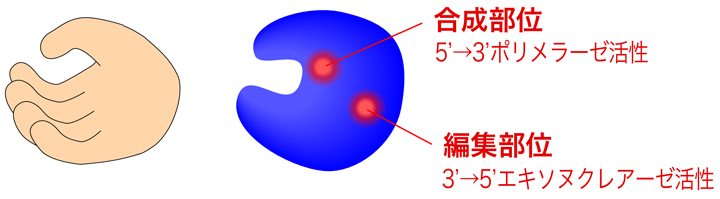
DNAポリメラーゼの構造は、よく右手に例えられる。この右手を開いたり閉じたりしながら、DNAを合成していく。実は、DNAポリメラーゼはDNAを合成するだけではなく、上の表にもあるようにDNAを端から削るエキソヌクレアーゼ活性ももつ。とくに多くのポリメラーゼがもつ3’→5’エキソヌクレアーゼ活性がDNAポリメラーゼによる正確な合成に重要となる。
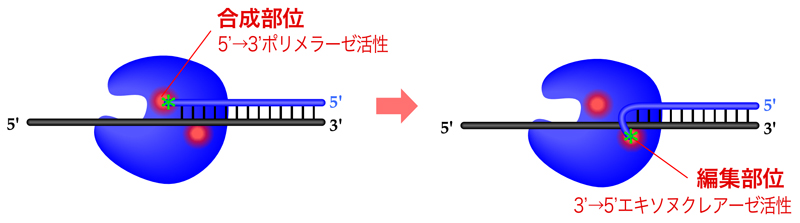
DNAポリメラーゼの3’→5’エキソヌクレアーゼ活性は、校正機能を担う。通常は合成部位の5’→3’ポリメラーゼ活性により鋳型と相補的なヌクレオチドを取り込んでいくが、105塩基に一回くらいの頻度で取り込みミスを犯す。誤ったヌクレオチドの取り込みによって3’末端が適切に塩基対を形成できてない状態では、DNAポリメラーゼは次のヌクレオチドを付加できない。そこで、編集部位の3’→5’エキソヌクレアーゼ活性により誤った塩基を端から削り、ミスのなくなった(3’末端が塩基対を形成している)ところで再び合成部位でのDNA合成を始める。こうした校正機能により、DNA合成の正確性は100倍上昇する。
原核生物のDNAポリメラーゼIのみがもつ5’→3’エキソヌクレアーゼ活性は、DNA合成の終了に関与するが、これはこのページの最後に説明したい。
複製フォークではたらく因子
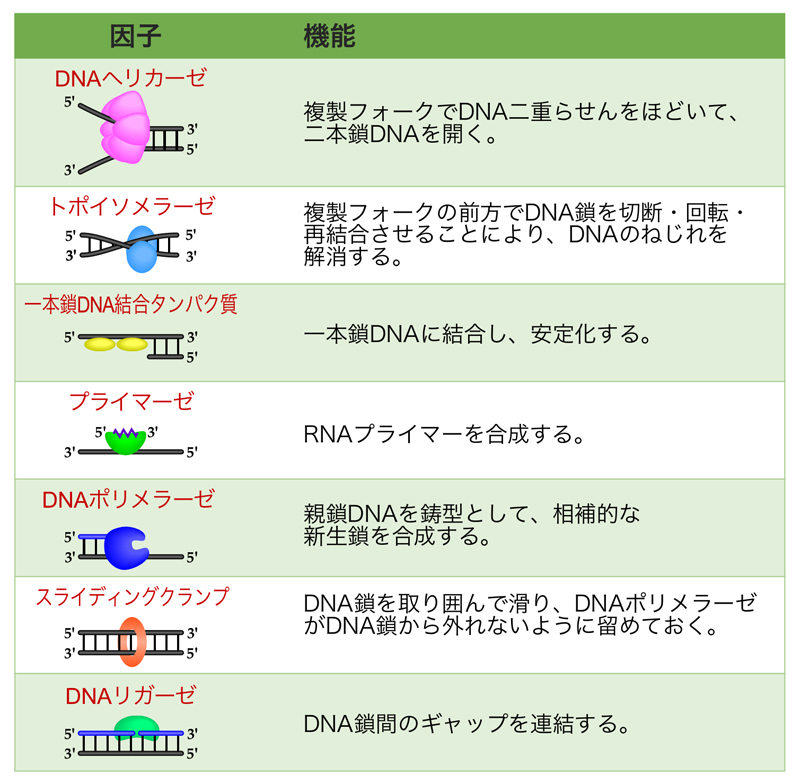
DNA複製は複製起点で始まり、そこから両方向にDNA合成が進行していく。このDNA合成を最先端で行っている現場が、複製フォークである。この複製フォークでは、どのような因子がどのように役割分担してDNA合成が行われているか、ここで説明していこう。要点は、右の表にまとめてある。ここでは、原核生物と真核生物の区別なく、共通にはたらく因子の概要のみ示している。
- DNAヘリカーゼ
DNAの二重らせんをほどいて一本鎖DNAにする酵素である。六量体でリング状の構造をとり、一本鎖DNAを取り囲むように結合する。6つのサブユニットが、ATPを加水分解しながら変形して元に戻る過程を順次繰り返しながらDNA鎖上を移動し、DNA二重らせんをほどいていく。DNA複製では、ラギング鎖の鋳型DNA上を5’→3’方向に移動し、DNA鎖をほどいていく。 - トポイソメラーゼ
DNAヘリカーゼがDNA二重らせんをほどいて開くと、その前方ではDNAが強く巻きすぎた状態になる。そこで、トポイソメラーゼがDNA鎖の強く巻きすぎた状態を解消する。Ⅰ型のトポイソメラーゼⅠは、DNAの一本鎖に切れ目(ニック)を入れ、切れ目の部分の5’末端(リン酸基)と一時的に共有結合する。この間に、切れ目のないDNA鎖を軸に切れ目の入った鎖を回転させた後、再びDNAの切れ目を連結させる。
Ⅱ型のトポイソメラーゼⅡは、2本のDNA鎖が交差するところで、DNA鎖どうしの絡みを解消する。すなわち、ATP加水分解のエネルギーを利用してDNAの両方の鎖を切断し、この切断部にもう一方のDNAを通過させ、再びDNA鎖を連結する。こうすることで、DNA複製後に染色体が絡まるのを防いでいる。 - 一本鎖DNA結合タンパク質
DNAヘリカーゼの作用で生じた一本鎖DNAに結合して一本鎖の状態を安定化する。すなわち、再び二本鎖DNAに戻るのを防ぐとともに、ヘアピン構造のようなDNA合成の障害となる二次構造の形成も防ぐ。一本鎖DNAの塩基部分を覆ってしまうことなく強く結合するので、一本鎖DNA結合タンパク質が結合したDNAはDNA合成の鋳型として機能できる。 - プライマーゼ
DNA合成のきっかけとなるプライマーを合成する。細胞内において、DNA複製に使われるプライマーは、約10ヌクレオチドのRNAである。転写を考えるとわかるが、RNAの合成は何もないところから始められるのだ。 - DNAポリメラーゼ
DNA合成を触媒する酵素。 - スライディングクランプ
日本語でいうと「滑る留め金」。DNAをリング状に取り囲んで、DNAポリメラーゼが鋳型鎖から離れないように支える環状のタンパク質複合体。スライディングクランプの装着には、クランプローダーというタンパク質複合体が必要である。 - DNAリガーゼ
DNA合成終了の際に、DNA鎖間のギャップを連結させる酵素。ラギング鎖では、岡崎フラグメント間の連結を行う。
複製フォークでのDNA合成
では、複製フォークにおけるDNA合成の機構を説明していこう。
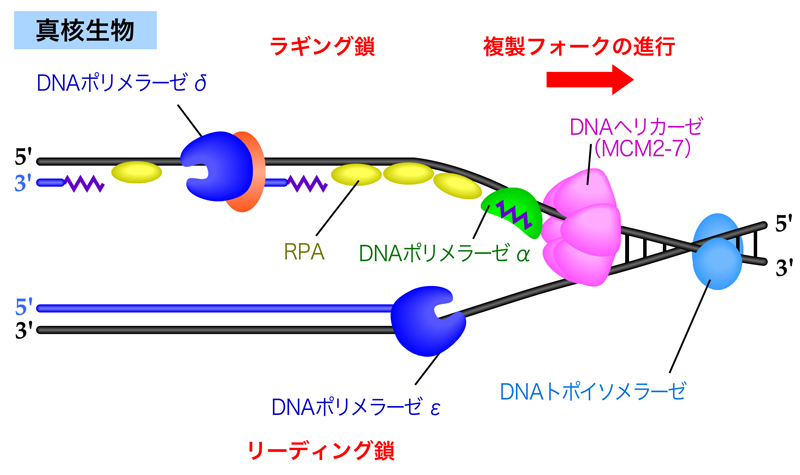
複製フォークの最先端部では、DNAヘリカーゼがDNA二重らせんをほどく。生じた一本鎖DNAには一本鎖結合タンパク質が結合し、二本鎖DNAに戻らないよう、またヘアピン構造をとらないよう一本鎖DNAの状態を安定化する。
リーディング鎖では、DNA合成開始のときにプライマーゼがRNAプライマーを合成し、そこから複製フォークの進行方向と同じ方向にDNAポリメラーゼが連続的に新生鎖を合成していく。新たに合成された二本鎖DNAを取り囲むようにスライディングクランプが結合しており、そこにDNAポリメラーゼが密着している。こうすることで、DNAポリメラーゼは合成中もDNAから外れずに保持され、しかもスムーズに移動できるのである。
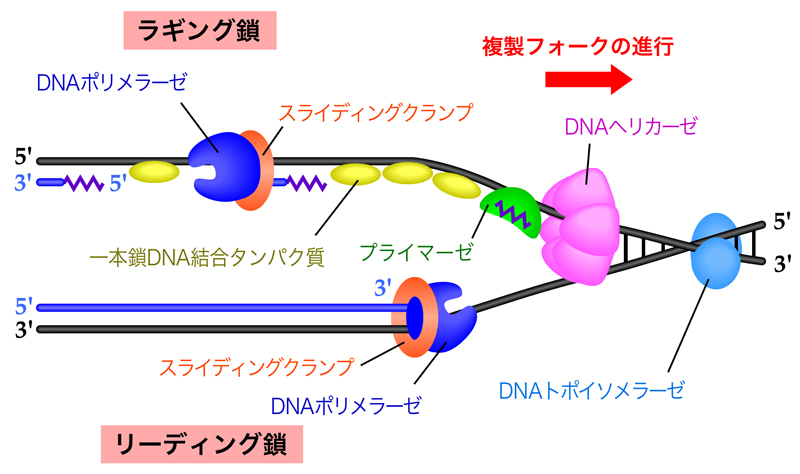
一方ラギング鎖では、プライマーゼによりRNAプライマーが合成された後に、DNAポリメラーゼが複製フォークの進行方向とは逆向きにDNA合成を行う。DNA合成が隣接する岡崎フラグメントに到達すると、DNAポリメラーゼは解離し、複製フォーク側に移動して次の岡崎フラグメントのDNA合成を始める。そして最後は、DNAリガーゼにより岡崎フラグメントが連結されて、ラギング鎖の合成が終了する。
原核生物と真核生物の違い
上記の説明は、原核生物/真核生物関係なく行われるDNA複製の概要である。原核生物と真核生物で概ね同じようにDNA複製は行われるが、詳細は色々と異なる。そこで、原核生物と真核生物の複製フォークでのDNA合成を比較してみよう。
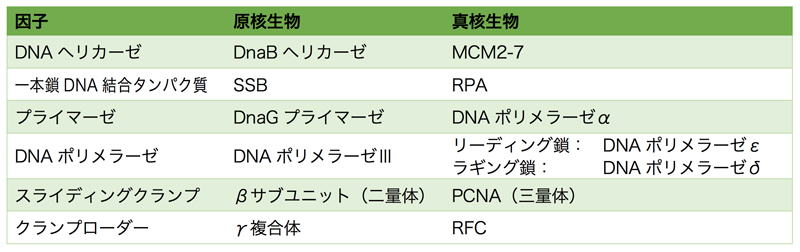
RNAプライマーの合成は、原核生物ではDnaGプライマーゼが行い、それに続いてDNAポリメラーゼⅢがDNA合成を開始する。一方真核生物では、RNAプライマーの合成はDNAポリメラーゼαにより行われる。DNAポリメラーゼαはプライマーゼと複合体を形成しており、このプライマーゼによりRNAプライマーが合成された後に、DNAポリメラーゼαが約30塩基のDNAを合成し、その後別のDNAポリメラーゼにスイッチして残りのDNAを合成する。
メインのDNA合成を行うのは、原核生物ではDNAポリメラーゼⅢがすべてを担当する。真核生物では、リーディング鎖とラギング鎖で異なるDNAポリメラーゼが担当すると考えられている。DNAポリメラーゼεは、スライディングクランプなどの補助因子なしでも高い伸長性をもつため、リーディング鎖の合成を担当する。DNAポリメラーゼδは、前方のDNA-RNA二本鎖部分を引き剝がしながら合成できることから、ラギング鎖の合成を担当する。
スライディングクランプは、原核生物ではDNAポリメラーゼⅢホロ酵素中のβサブユニットの二量体。真核生物では、PCNAの三量体である。二量体と三量体の違いはあるものの、どちらも似たようなリング上の構造を形成する。クランプローダーも、原核生物ではDNAポリメラーゼⅢホロ酵素中のγ複合体、真核生物ではRFCで、どちらも5つのサブユニットで構成される。
DNA合成の終了
ラギング鎖のDNA合成が隣接するラギング鎖に到達するとDNA合成は終了だが、ではどのように終了するのだろうか。
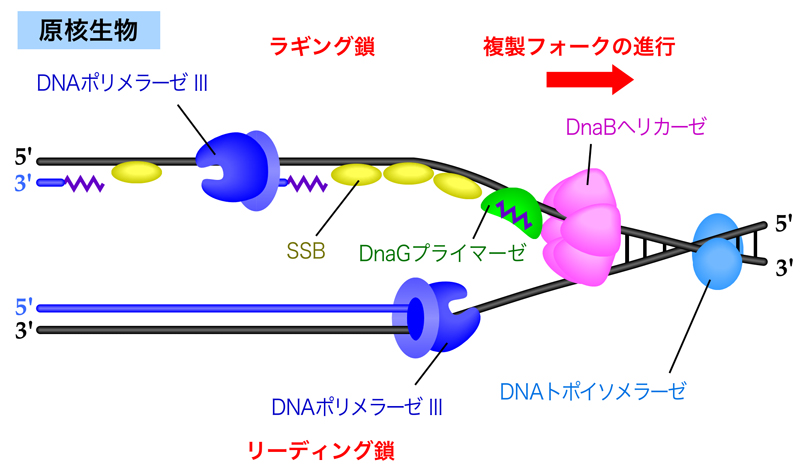
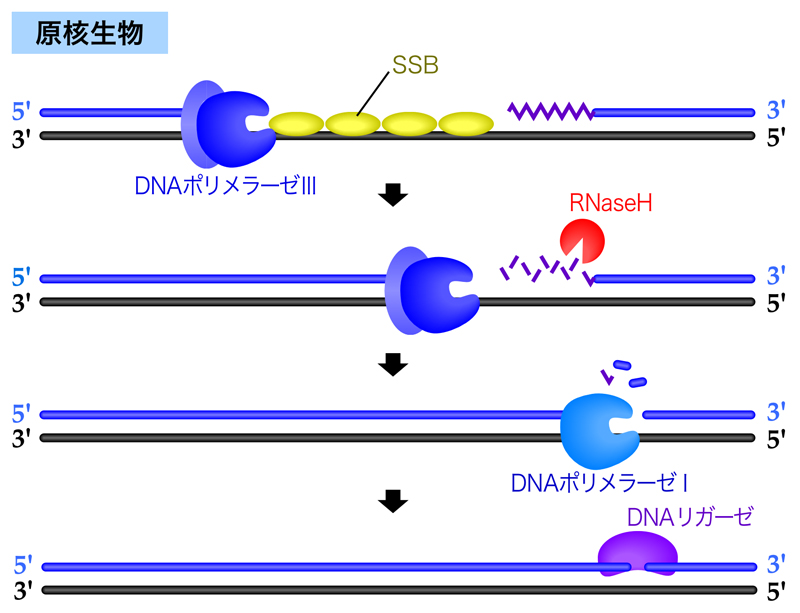
まずは原核生物を例に説明しよう。DNAポリメラーゼⅢによるDNA合成が隣接するラギング鎖に到達すると、役目を終えて離れてしまう。ここで、RNAプライマーを除去するために、DNA-RNAハイブリッド中のRNAを分解するRNaseHがRNAプライマーを分解する。しかし、RNaseHはDNAに結合した最後のRNAヌクレオチドは分解できない。そこで、DNAポリメラーゼⅠがその5’→3’エキソヌクレアーゼ活性により、前方の岡崎フラグメントをある程度削り込みながら、DNAを合成していく。そして最後は、DNAリガーゼが2つの岡崎フラグメントを連結する。
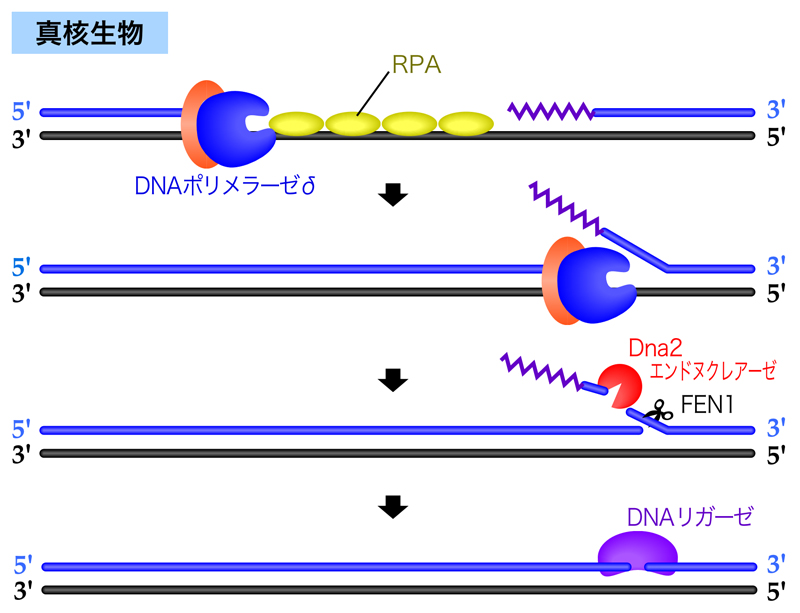
真核生物では、ラギング鎖の合成はDNAポリメラーゼδが行う。しかし、DNAポリメラーゼδには5’→3’エキソヌクレアーゼ活性がないので、原核生物とは異なる方法でRNAプライマーの除去を行う。DNAポリメラーゼδが隣接するラギング鎖に到達すると、前方のRNAプライマーを引き剝がしながらDNA合成を継続する。DNAポリメラーゼαは3’→5’エキソヌクレアーゼ活性(校正機能)をもたないため、RNAプライマーに続く領域のDNA合成も決して正確ではない。そこで、DNAポリメラーゼδはDNAポリメラーゼαによって合成された領域も一気に引き剝がし、この引き剥がされた鎖をDna2エンドヌクレアーゼやFEN1などのヌクレアーゼが切断・除去し、最後はDNAリガーゼが2つの岡崎フラグメントを連結する。
リーディング鎖とラギング鎖の協調
ここまで、複製フォークで行なわれているDNA合成の分子メカニズムを説明してきたが、各因子はバラバラに役割分担をして機能を果たしているわけではない。DNAポリメラーゼにしても、リーディング鎖とラギング鎖でバラバラに逆方向にDNA合成を行っていたのでは、効率が悪いだろう。やはり、効率よくDNA複製を行うためには、これらの因子が近くに集まって協調的に進めるのがよいだろう。
そこで、下図のようにラギング鎖のDNAを折り返して、リーディング鎖のDNAポリメラーゼとラギング鎖のDNAポリメラーゼが複合体を形成し、同じ方向にDNA合成できるようにしていると考えられている。しかもこうすると、岡崎フラグメントのDNA合成を終えたときに、次に合成すべき岡崎フラグメントの合成開始部位(RNAプライマー)がすぐ近くに位置することになり、DNAポリメラーゼはスムーズに次のDNA合成に移ることができる。このようなモデルは、トロンボーンのスライド管のように伸び縮みすることからトロンボーンモデルとよばれる。