原核生物の転写調節の2ページ目です。ここでは、調節タンパク質を介した転写調節のしくみを紹介しよう。調節タンパク質を介したしくみは、真核生物におけるしくみとも共通する部分があります。そこで、Lacオペロンを例に、調節タンパク質を介した調節機構について紹介します。
転写調節タンパク質
原核生物の転写調節は、RNAポリメラーゼのσ因子を切り替えることでも行われるが、調節タンパク質を介した調節も主要な調節機構である。調節タンパク質というのは、RNAポリメラーゼとは別に、転写を活性化あるいは抑制するタンパク質のことである。このような調節タンパク質は、遺伝子の外部から作用するので、トランス作用性因子ともよばれる。もちろん、調節タンパク質も遺伝子によってコードされている。
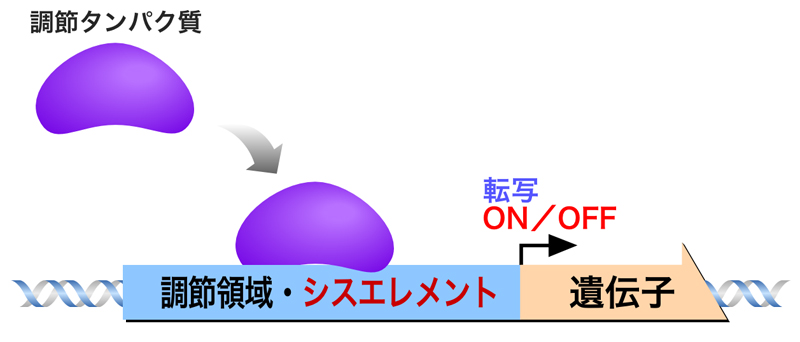
調節タンパク質は、遺伝子の転写調節領域にある標的配列に結合する。このような標的配列は遺伝子と同じDNA分子内にあるため、シスエレメントとよばれる。
正の調節と負の調節
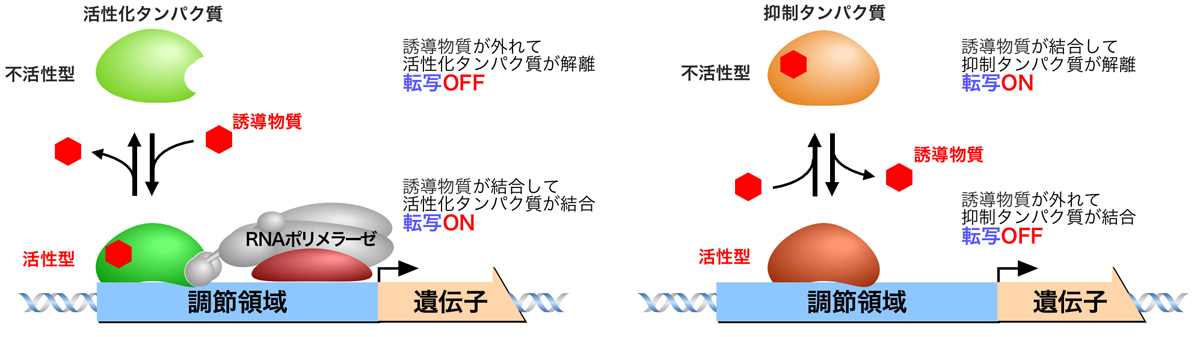
調節タンパク質を介した転写調節には、2つのモードがある。
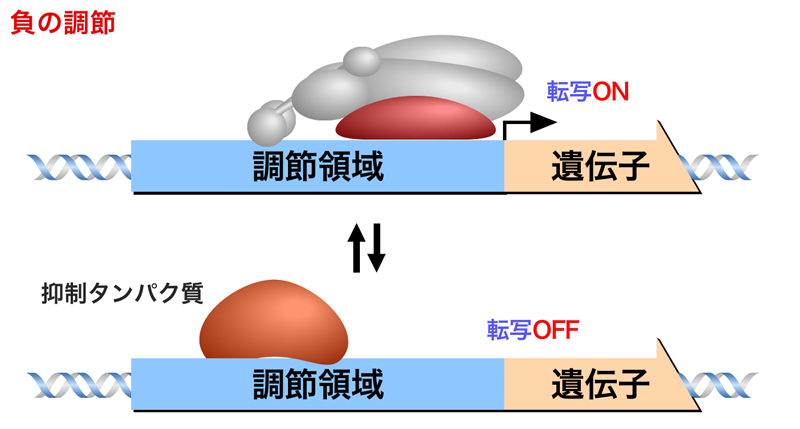
負の調節は抑制タンパク質を介した調節であり、抑制タンパク質がシスエレメントに結合すると転写が抑制されるが、抑制タンパク質が離れることにより遺伝子の転写がONになる。
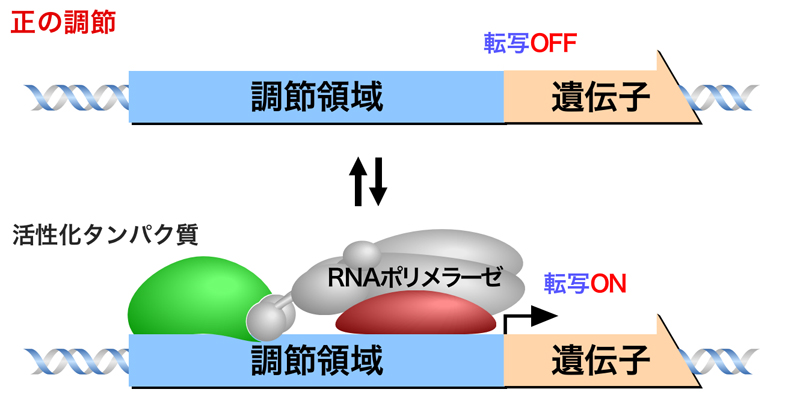
正の調節は活性化タンパク質を介した調節であり、活性化タンパク質がシスエレメントに結合することにより遺伝子の転写がONになる。
原核生物は、生育環境の変化に迅速に対応するとともに、不必要なものにはエネルギーを使わないような効率の良さをもっている。例えば、酵素の基質が存在しないときにはその酵素の合成をストップし、基質が供給されるとすぐに対応して酵素を合成する。また、酵素による反応産物が過剰にあるときには酵素の合成をストップし、反応産物が欠乏すると酵素を合成する。このような対応に、負の調節と正の調節が使われる。
酵素の基質が存在しないときは、抑制タンパク質がシスエレメントに結合して酵素遺伝子の転写を抑制するが、基質が供給されると基質が誘導物質(inducer)となって抑制タンパク質に結合し、抑制タンパク質による抑制が解除されて転写がONになる(負の調節)。あるいは、基質が誘導物質となって活性化タンパク質に結合し、これがシスエレメントに結合して酵素遺伝子の転写がONになる(正の調節)。
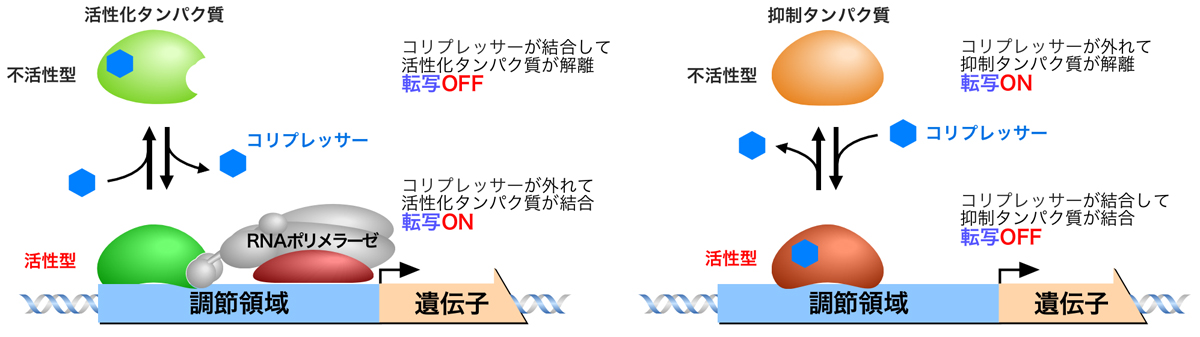
また、酵素の反応産物が欠乏しているときは活性化タンパク質がシスエレメントに結合して酵素遺伝子の転写をONにしているが、過剰なときは反応産物がコリプレッサー(corepressor)となって活性化タンパク質に結合し、これがシスエレメントに結合して酵素遺伝子の合成をOFFにする(正の調節)。あるいは、反応産物がコリプレッサーとなって抑制タンパク質に結合し、これがシスエレメントに結合して酵素遺伝子の合成をOFFにする(負の調節)。
Lacオペロン
ではここで、上記の負の調節と正の調節についてLacオペロンを例に説明しよう。
オペロンとは、1つのオペレーターで制御される転写単位とその制御エレメントのセットのことである。そしてこのオペレーターが、オペロンの転写を起こさせるか否かを決定するシスエレメントである。
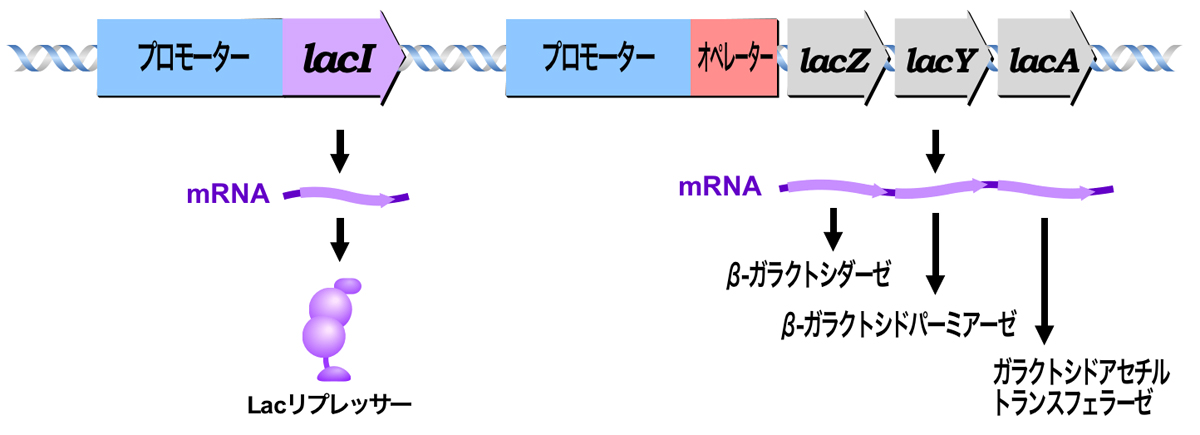
Lacオペロンはラクトースの代謝系を制御するオペロンであり、ラクトースの代謝に関わる酵素群をコードするlacZ, lacY, lacA遺伝子の転写が制御される。lacZ遺伝子はβガラクトシドを単糖に分解する酵素βガラクトシダーゼをコードし、lacY遺伝子はβガラクトシドを細胞内に取り込む酵素βガラクトシドパーミアーゼをコードし、lacA遺伝子はβガラクトシドにアセチル基を転移する酵素ガラクトシドアセチルトランスフェラーゼをコードする。
lacZ, lacY, lacA遺伝子は、1つのプロモーターから転写される1つの転写単位である。この遺伝子群の転写は、プロモーター近傍のオペレーターと、隣にあるlacI遺伝子産物により制御される。lacI遺伝子は抑制タンパク質であるLacリプレッサーをコードしており、lacZ, lacY, lacA構造遺伝子とは別のプロモーターから転写される独立の転写単位である。
Lacオペロンにおける負の調節
ラクトースが存在しないとき、Lacリプレッサーは四量体を形成し、Lacオペロンのオペレーターに結合する。Lacリプレッサーの結合によりRNAポリメラーゼによる転写が妨げられ、lacZ, lacY, lacA構造遺伝子の発現はOFFになる。
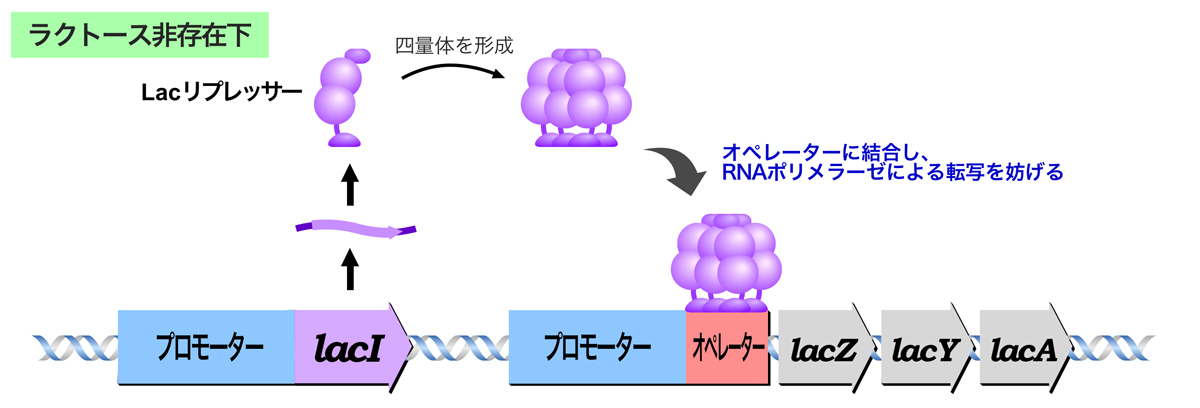
Lacリプレッサーのオペレーターへの結合が転写を妨げるしくみについては諸説ある。1つは、Lacリプレッサーのオペレーターへの結合がRNAポリメラーゼのプロモーターへの結合と競合し、RNAポリメラーゼがプロモーターに結合できないという説。もう一つは、Lacリプレッサーのオペレーターへの結合はRNAポリメラーゼのプロモーターへの結合と同時に起こるが、このときRNAポリメラーゼが転写を開始できないという説。詳細は、また別のページで紹介しよう。
ラクトースが生育環境に存在すると、ラクトースが誘導物質となってLacリプレッサーに結合し、Lacリプレッサーの構造が変化してオペレーターに結合できなくなるため、Lacリプレッサーによる抑制が解除され、lacZ, lacY, lacA構造遺伝子の発現がONになる。
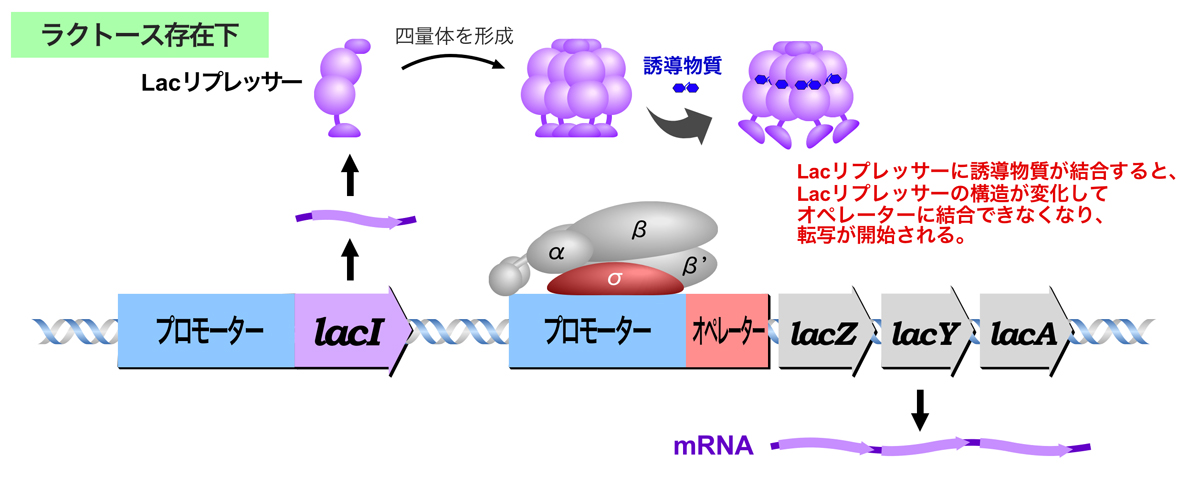
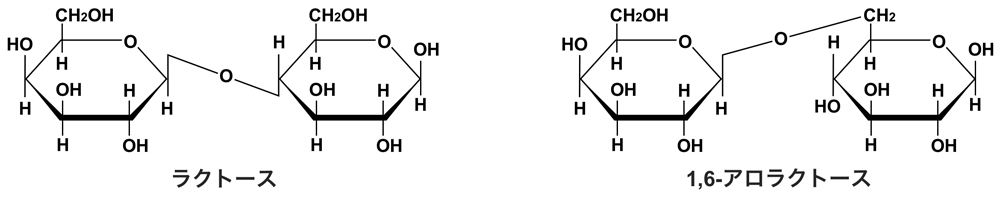
Lacリプレッサーによる抑制解除の生理的な誘導物資は1,6-アロラクトースである。1,6-アロラクトースは、lacZ遺伝子にコードされるβ-ガラクトシダーゼによるラクトースの糖転移によって生成する。ラクトースがない状態でもlacZ, lacY, lacA構造遺伝子が基底レベル(大腸菌内に数分子程度)で発現しているから、ラクトース添加後すぐに対応できるようである。
Lacオペロンにおける正の調節
Lacオペロンの発現には、正の調節も関与する。
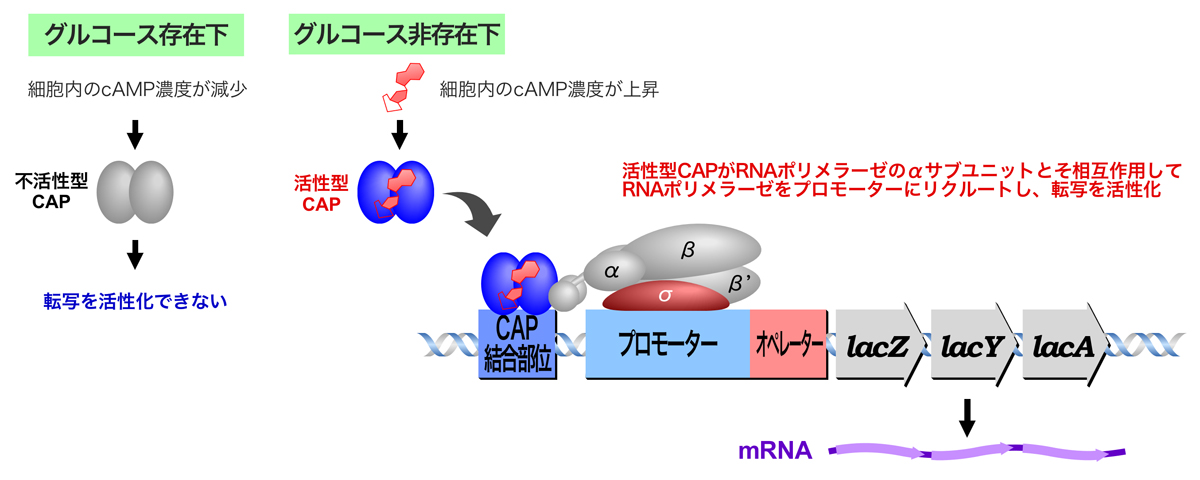
大腸菌は、生育環境中にグルコースが存在すると、エネルギー源として効率のよいグルースを優先的に利用し、他の糖類の代謝に関わる遺伝子の発現を抑制する。このような現象をカタボライト抑制(異化代謝産物抑制)といい、無駄な酵素産生を防いでいる。
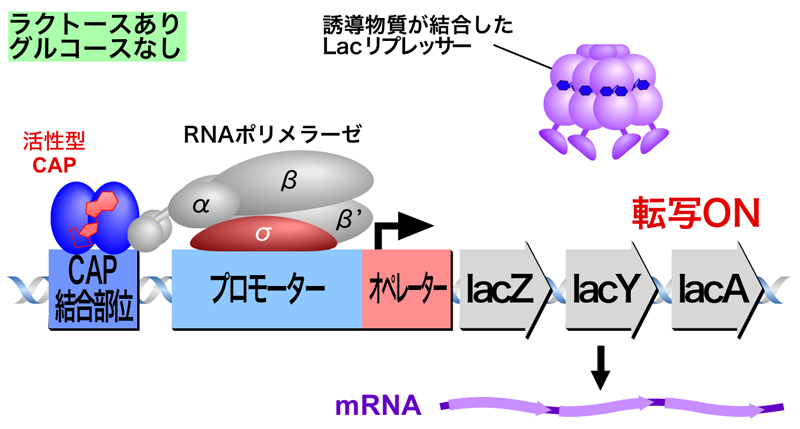
生育環境中にグルコースがあると、細胞内のcAMP濃度は減少している。しかし、生育環境中にグルコースが存在しないと細胞内のcAMP濃度が上昇し、cAMPが結合したカタボライト活性化タンパク質CAP(またはcAMP受容タンパク質CRPともよばれる)が活性型に変化する。活性型CAPはプロモーター近傍のCAP結合部位に結合するとともに、RNAポリメラーゼのαサブユニットのC末端ドメイン(CTD)と直接相互作用することにより、RNAポリメラーゼを効率よくプロモータ上にリクルートし、Lacオペロンの転写を活性化する。
Lacオペロンにおける転写調節
Lacオペロンの転写は、上記の負の調節と正の調節の両者により決定される。
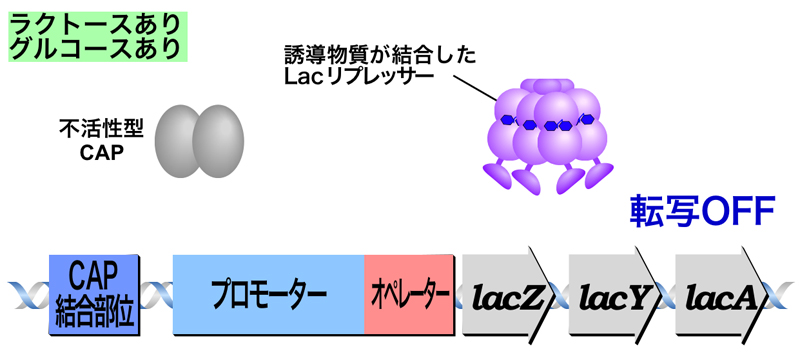
- 生育環境中にラクトースとグルコースが両方あるとき
ラクトースの存在によりLacリプレッサーによる抑制は解除されるが、グルコースの存在によりCAPが活性化されないため、転写はOFFのままである。
- 生育環境中にラクトースとグルコースがどちらも存在しないとき
グルコースが存在しないためCAPが活性化されてCAP結合部位に結合するが、ラクトースが存在しないためLacリプレッサー四量体がオペレーターに結合して転写が抑制され、転写はOFFのままである。

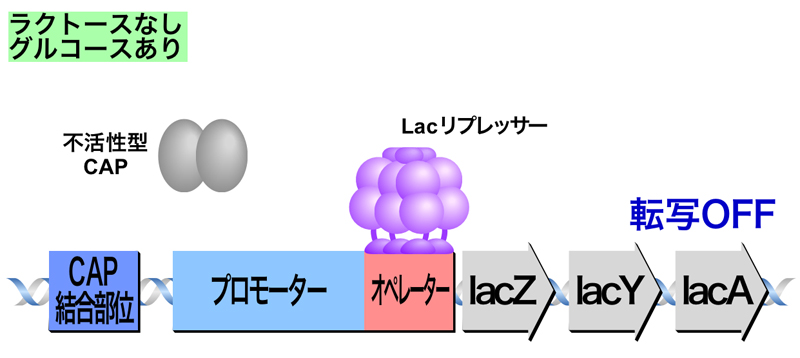
- 生育環境中にラクトースは存在せず、グルコースがあるとき
ラクトースの存在しないためLacリプレッサー四量体がオペレーターに結合して転写が抑制されており、さらにグルコースの存在によりCAPが活性化されないため、転写はOFFのままである。
- 生育環境中にラクトースが存在するが、グルコースは存在しないとき
ラクトースの存在によりLacリプレッサーによる抑制は解除され、さらにグルコースが存在しなためにCAPが活性化されてCAP結合部位に結合するため、RNAポリメラーゼが効率よくプロモーター上にリクルートされて転写はONになる。