続いて、転写調節のメカニズムについて紹介していきましょう。原核生物の転写調節から入りますが、この頁では、σ因子を介した転写調節のしくみについて説明します。ただし、ここでいう原核生物とは細菌(真正細菌)のことなので、注意しよう。
原核生物の転写
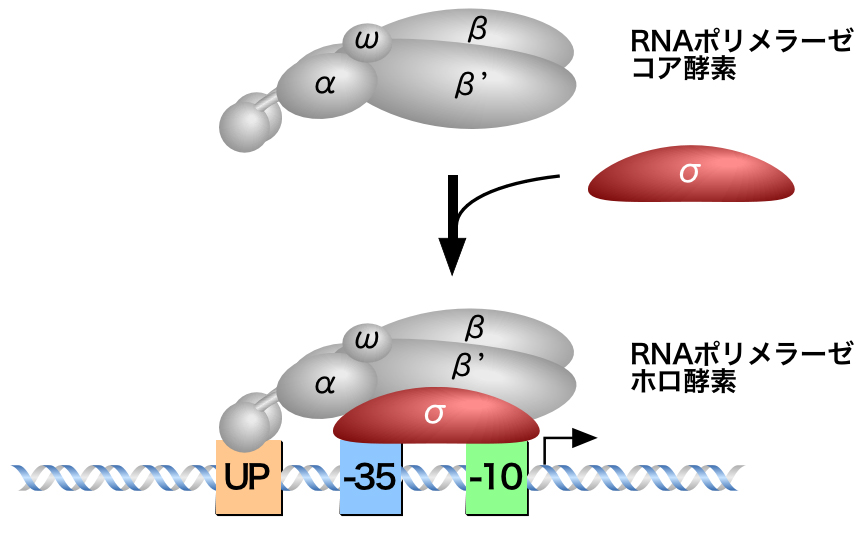
原核生物は、1種類のRNAポリメラーゼがすべての遺伝子の転写を行う。2つのαサブユニットとβ・β’・ωサブユニットが1つずつ集まってRNAポリメラーゼコア酵素が形成されるが、このコア酵素はプロモーターを特異的に認識できない。
そこで、プロモーターへの特異的結合を担うσサブユニット(σ因子)がコア酵素に結合することで、RNAポリメラーゼはホロ酵素となり、RNAポリメラーゼにプロモーターへの特異性が付与される。したがって、RNAポリメラーゼホロ酵素はプロモーターを特異的に認識して結合し、特定の遺伝子の転写を活性化することができる。
σ因子を交換する
原核生物には、このσ因子を介した転写調節のしくみがある。すなわち、RNAポリメラーゼのσ因子を、プロモーターへの特異性が異なる別のσ因子取り替えることによって、転写される遺伝子群を切り替えるのである。
そこでよく例に出されるのが、熱ショック遺伝子の転写調節である。大腸菌では通常、σ70というσ因子をもつRNAポリメラーゼによって大部分の遺伝子が転写される。ところが大腸菌が熱ショックにさらされると、細胞を熱ショックから守るための遺伝子が転写されるのだが、このときに使われるのがσ32という別のσ因子である。
σ32遺伝子のプロモーターも、σ70をもつRNAポリメラーゼによって転写されるが、大腸菌が熱ショックにさらされると細胞内におけるσ32の量が増加する。そして、一部のRNAポリメラーゼでσ70に代わってσ32が取り込まれ、このσ32をもつRNAポリメラーゼが熱ショック遺伝子の転写を活性化させるのである。

では、なぜ熱ショックによりσ32の量が増加するのだろうか。実は、σ32の翻訳効率が上昇するのである。σ32 mRNAは、σ70をもつRNAポリメラーゼによって転写されている。しかし通常の生育温度では、mRNAが分子内塩基対を形成することにより立体的に折りたたまれているために、翻訳開始に必要なリボソーム結合部位(シャインダルガーノ配列:SD配列)が利用できない。ところが熱ショックにさらされると、σ32 mRNAの立体構造が壊れてSD配列を利用できるようになり、翻訳効率は上昇するのである。

大腸菌のσ因子
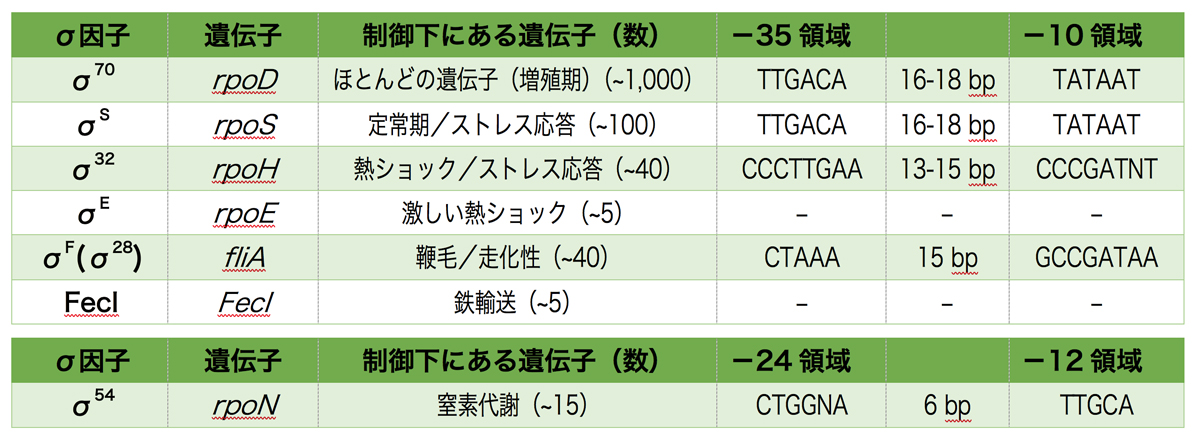
σ因子にはσ70型因子とσ54型因子があり、それぞれが特定の遺伝子の転写に関わる。ほとんどの原核生物は複数のσ70型のσ因子をもつが、σ54型についてはあっても1つ以上はない。したがって、σ70型とσ54型の両方をもつ細菌もいれば、σ70型のみの原核生物もいる。σ70型の数についてもさまざまで、マイコプラズマ(Mycoplasma sp.)のσ70型は1個だけだが、放線菌(S. coelicolor)のσ70型は60個以上ある。
では、大腸菌を例に紹介しよう。大腸菌は下表のように、6種類のσ70型因子と1種類のσ54型因子をもつ。
- σ70は増殖に発現するほとんどのハウスキーピング遺伝子の転写を担う。
- σSは定常期における転写を担うσ因子で、結合するプロモーター配列(-35と-10領域)はσ70と一緒であるが、プロモーターの選択には調節タンパク質も関与する。σSはさまざまなストレスシグナルで活性化され、ストレス応答に関連する遺伝子の転写を担う。
- σ32は熱ショックや他のストレスによって生じた細胞質中の変性タンパク質により誘導され、変性タンパク質を正しくフォールディングしたり分解したりする遺伝子を発現させる。
- σEはもっと激しい温度変化に応答する遺伝子の転写を担う。細胞膜の変性したタンパク質によって誘導され、膜を元に戻すのに必要な遺伝子を発現させる。
- σ28(σF)は走化性や鞭毛形成に関わる遺伝子の転写を担う。
- FecIは、鉄欠乏時にクエン酸鉄を細胞内に取り込むための輸送装置をコードするfecABCDEオペロンの転写を担う。
- σ54型因子であるσ54は、培地のアンモニアが欠乏したとき(窒素飢餓条件下)に、窒素代謝に関する遺伝子の転写を担う。
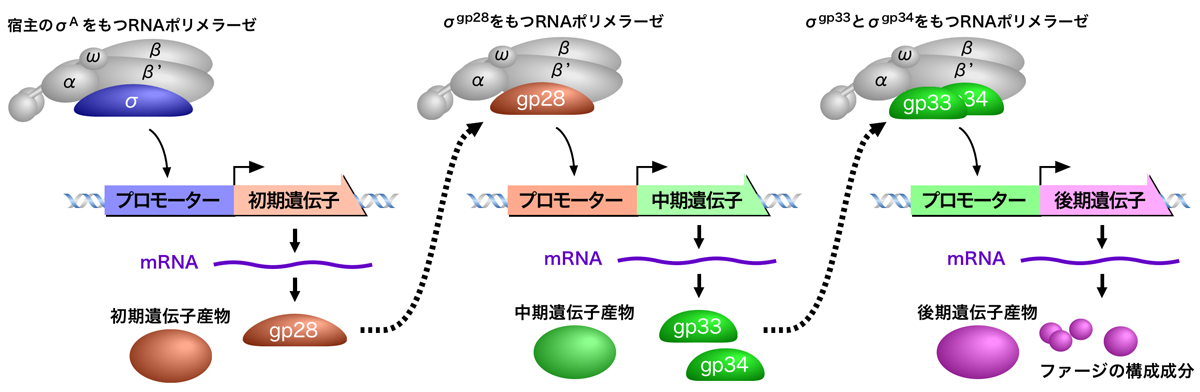
ファージにおけるσ因子を介した制御
σ因子はRNAポリメラーゼがどのプロモーターから転写を開始するか、その特異性を決める。上記の大腸菌と同様、枯草菌(Bacillus subtilis)も複数(10種類)のσ因子をもっており、栄養源が枯渇して飢餓状態になったときに増殖を停止して休止状態の胞子を形成したり、大幅な代謝の変化をもたらす場合に利用される。
枯草菌では、主要なσ因子はσAである(もともとSDS-PAGEにより推定される分子量からσ55とよばれ、その後クローニングされた遺伝子の配列から推定される分子量によりσ43とよばれたが、混乱を避けるためにアルファベットによる命名が使われるようになった)。しかし、熱ショックや浸透圧ショックなどの一般的なストレスへの応答にはσB(σ37)が利用され、胞子形成が始まるとσE(σ29)が利用される。
このようなσ因子の交換による遺伝子発現の切り替えは、ファージ(細菌に感染するウイルス)が感染後に自身の遺伝子を転写させるのにも使われる。つまり、ファージは自身のゲノムを転写するのに必要なσ因子をもっているのである。ここでは、枯草菌に感染するSPO1ファージを例に説明する。
SPO1ファージが宿主の枯草菌に感染すると、宿主のσAをもつRNAポリメラーゼによりSPO1ファージの初期遺伝子が転写される(枯草菌が胞子形成に入ってσAが欠損すると、SPO1ファージは感染しても増殖できない)。この初期遺伝子には、次の中期遺伝子の発現に必なgp28というσ因子(σgp28)をコードするものがある。このσgp28が、宿主のRNAポリメラーゼがもつσAと入れ替わることにより、今度はSPO1ファージの中期遺伝子が転写される。中期遺伝子には、後期遺伝子の発現に必須のgp33とgp34というσ因子(σgp33とσgp34)をコードするものがあり、これらがRNAポリメラーゼに取り込まれてSPO1ファージの後期遺伝子が発現するようになる。後期遺伝子にはファージの構成成分がコードされており、これにより子孫ファージが構築されて増殖していくのである。