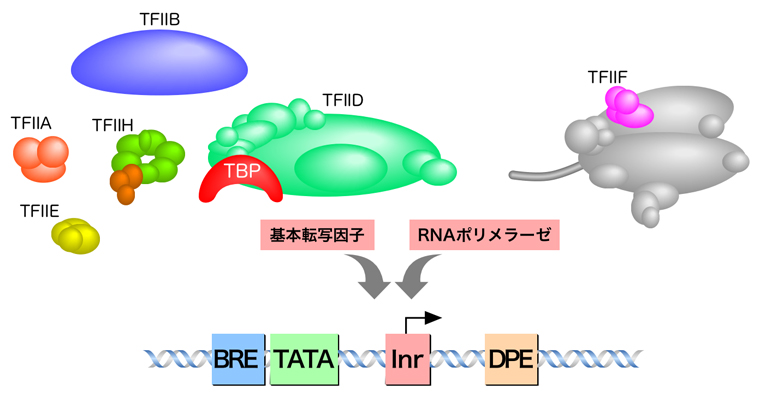
真核生物のRNAポリメラーゼは、正確な転写開始のために基本転写因子を必要とする。このページでは、基本転写因子について少し解説しよう。
基本転写因子
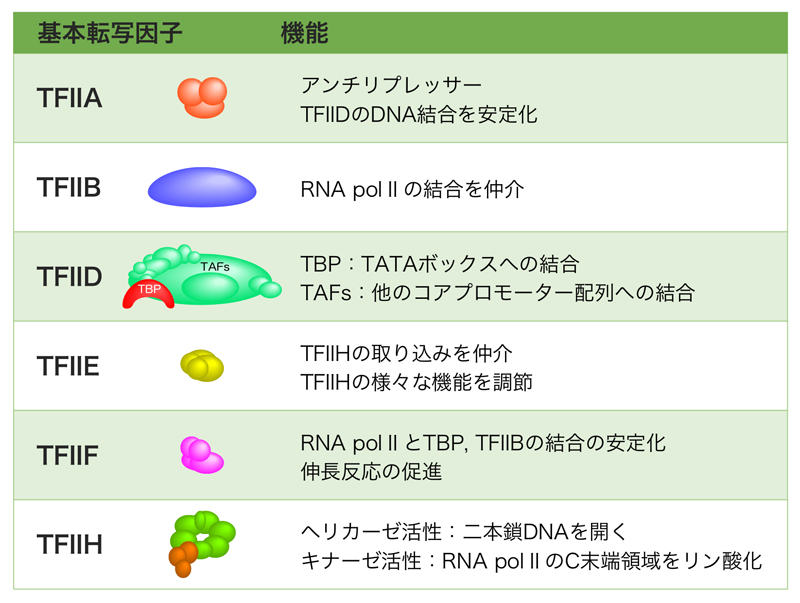
基本転写因子は、RNAポリメラーゼが正しくプロモーターを認識して転写開始するのに必要な因子である。RNAポリメラーゼIIによる転写に必要な基本転写因子は(TFIIA, TFIIB, TFIID, TFIIE, TFIIF, TFIIH)の6種類だが、その名称はTranscription Factor for RNA polymerase IIの頭文字をとり、そこにABCのアルファベットを割り振ったものである。その機能の概要は、下の表に示したとおりだが、詳細はさらに下に示す。
TFIID
TFIIDは、RNAポリメラーゼIIの基本転写因子の中でもとくに重要な役割を果たし、コアプロモーター配列中に存在するTATAボックス, Inr, DPE, DCE, MTEはすべてTFIIDにより認識される。TFIIDは、1分子のTATA結合タンパク質(TATA-binding protein:TBP)とおよそ10数種類のTBP関連因子(TBP-associated factor:TAF)により構成される。
TBPはその名のとおり、TATAボックスに結合するサブユニットである。生物種によって異なる構造をもつN端可変領域と、種間でよく保存されているC端保存領域がある。C端保存領域にはダイレクトリピートがあり、各繰り返し部位で2個のαヘリックスと5個の逆平行βシートを形成し、各ユニットが点対称に向き合うことで鞍型構造が形成される。
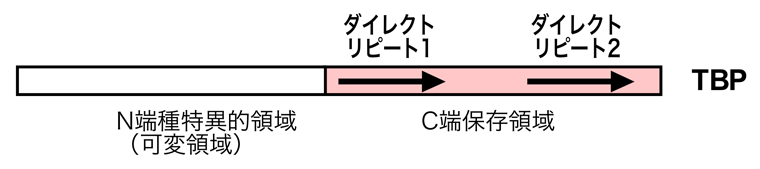

鞍型構造の内側のくぼんだ部分がDNAと接触し、DNA二重らせんの副溝(Minor Groove)を認識して結合する。TBPがDNAに結合すると、DNAが約80°曲がる。TBPには関連するファミリーが存在するが、それについてはまたどこかで紹介したい。
もう一つひとつの構成因子TAFだが、分離の方法や組織によってバリエーションがあり、構成が定まっているわけではない。以前は同じ因子がヒトではTAFII250、ショウジョウバエではTAFII230といったように、生物種によって異なる名称が与えられていたが、最近はTAF1〜TAF13といった統一的な名称がある。この中でも、最大のTAF1はTBPやいくつかのTAFと直接結合してTFIIDの核となるほか、ヒストンアセチル化活性・キナーゼ活性などももつ。TAFはコアプロモーターのコンセンサス配列も認識して結合する。例えば、TAF2はイニシエーター(Inr)に結合し、TAF6とTAF9は下流コアプロモーター配列(DPE)に結合する。
TFIIA
ヒトでは、TFIIAα, TFIIAβ, TFIIAγの3つのサブユニットがあり、αとβは1つの遺伝子がコードする前駆体タンパク質の切断により生じる。酵母では、TOA1とTOA2の2つのサブユニットがあり、TOA1のN末端・C末端はそれぞれ、ヒトのα・βサブユニットと相同性が高い。
TFIIAの機能は、TFIIDのDNAへの結合を促進・安定化するが、もう一つの重要な機能として、TBP特異的リプレッサーの機能を解除するアンチリプレッサー能をもつ。例えば、TFIIDに含まれるTAF1のTAND (TAF N-terminal domain)はTBPのDNA接触面を塞いで結合を阻害するが、TFIIAはこれを解除することができる。
TFIIB
TFIIBは、基本転写因子の中で唯一、単一サブユニットから構成される。TFIIDの次にプロモーターに結合する基本転写因子であり、PolⅡとTFIIDの橋渡しの役割を担う。
TFIIE
TFIIEαとTFIIEβがそれぞれ2つずつ結合したヘテロ四量体。先にDNAに結合しているTFIIB, TFIIF, PolポリメラーゼⅡと結合してTFIIHの取り込みを仲介し、TFIIHの様々な機能を調節する。
TFIIF
αサブユニット(RAP74)とβサブユニット(RAP30)によるα2β2のヘテロ四量体を形成している。TFIIB, RNAポリメラーゼⅡと結合することから、RNAポリメラーゼIIの呼び込みに関与すると考えられる。また、伸長反応中もRNAポリメラーゼIIに結合しており、伸長反応を促進する。
TFIIH
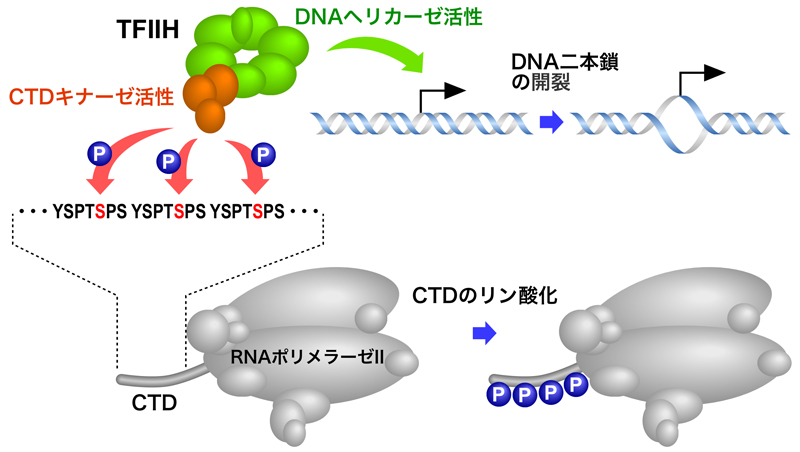
TFIIHは10個のサブユニットで構成され、そのうちの7個はリング状の構造をとってATP依存性DNAヘリカーゼ活性をもつ。DNAはこのリング状構造の中央の穴を通り、両側に位置するXPBとXPDが転写開始点上流10 bp付近からプロモーター部分のDNAを開裂する。
残りの3個(CAKサブコンプレックス)は球状構造をとり、CTDキナーゼ活性をもつ。RNAポリメラーゼのRpb1サブユニットのC末端ドメイン(CTD)は、YSPTSPSの7アミノ酸の反復構造で構成されており、その反復回数はヒトで52回、ショウジョウバエで43回、出芽酵母で26回となっている。TFIIHのCTDキナーゼ活性により、YSPTSPS配列中の5番目のS(セリン)がリン酸化され、このリン酸化CTDにRNAキャッピング酵素が結合する。
このように、DNAヘリカーゼ活性により二本鎖DNAを開き、CTDキナーゼ活性によりRNAポリメラーゼⅡのCTDをリン酸化することにより、転写が開始される。
転写開始前複合体の形成
以上の基本転写因子とRNAポリメラーゼIIがプロモーター上に集合することにより、転写開始前複合体が形成され、RNAポリメラーゼIIがプロモーター上の正しい位置に配置されることにより、正しい位置からの転写開始が実現されるのである。基本転写因子がプロモーター上に集合するプロセスについては、次のページで説明しよう。