
真核生物のRNAポリメラーゼIIによって合成されたmRNA前駆体は、細胞質へ運ばれる前にRNAプロセシングを受けて成熟mRNAとなる。このページでは、RNAプロセシングの全容を紹介しましょう。
原核生物と真核生物
原核生物には核膜で囲われた核はなく、転写の場と翻訳の場を仕切るものがない。したがって、転写によって合成されたmRNAは、すぐその場で翻訳されることになる。このように、原核生物では転写と翻訳が密接に連携している。
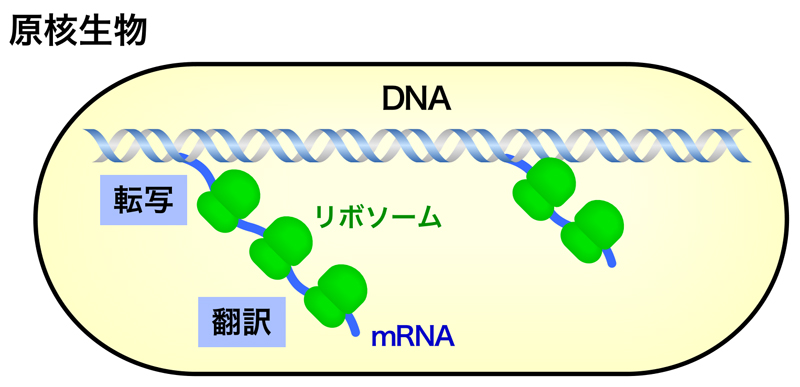
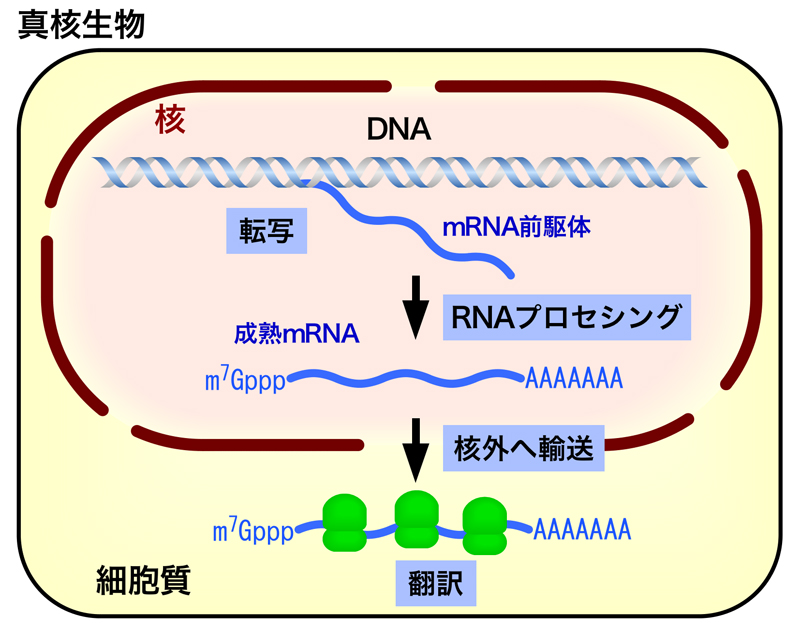
真核生物では、遺伝情報をもつDNAは核膜で囲われた核という構造体の中にあり、転写(RNAの合成)は核の中で行われる。一方、翻訳(タンパク質の合成)は、細胞質のリボソームで行われ、転写の場と翻訳の場は完全に仕切られている。したがって、核内でRNAポリメラーゼIIによって合成されたRNA(mRNA前駆体)は、翻訳の前に核外へと輸送されなくてはならない。この核外への輸送の前に、mRNA前駆体は様々な加工を受けて、成熟mRNAとなるのだが、この段階がRNAプロセシングとよばれる。RNAプロセシングには、キャップ構造の形成・RNAスプライシング・ポリA尾部の形成の3つの過程がある。
5’キャップ構造の形成
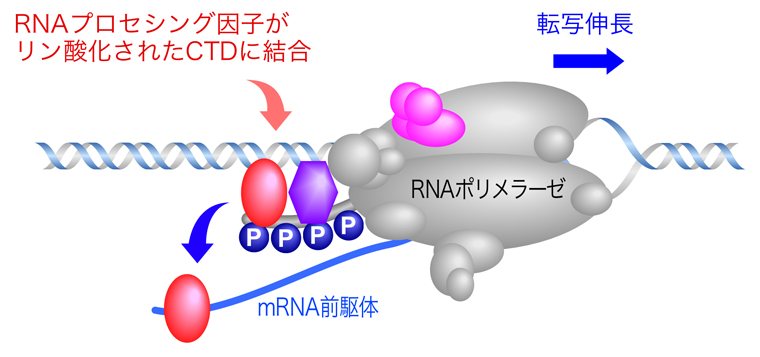
前の頁で説明したように、転写の伸長とRNAプロセシングとが共役している。リン酸化されたRNAポリメラーゼIIのRpb1サブユニットのCTDに、RNAプロセシング因子が結合しており、必要に応じて転写中のmRNA前駆体へ移動し、RNAプロセシングを実行するのである。しかも、プロモーター付近では5番目のセリン(Ser5)のリン酸化が高く、3’末端側に向かってSer5のリン酸化は低下し、代わりに2番目のセリン(Ser2)のリン酸化が高くなっている。
ここで、5’キャップ構造の形成について説明しよう。
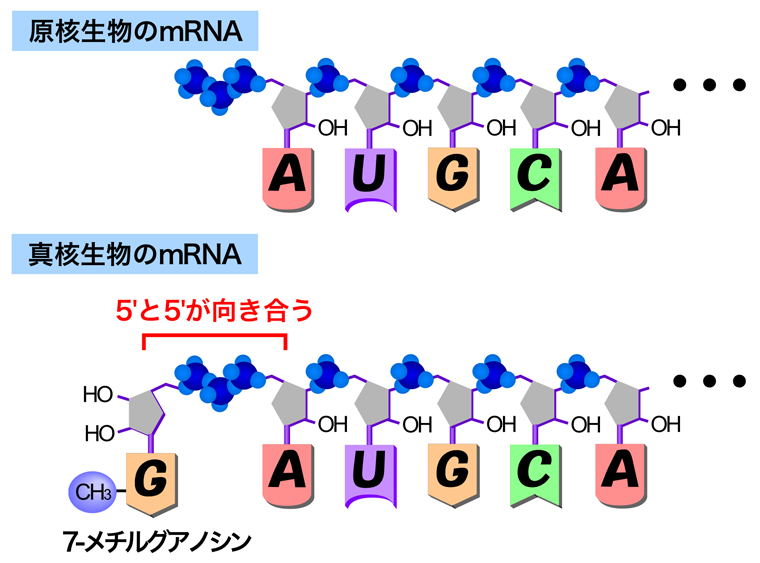
はじめに、原核生物と真核生物のmRNAの5’末端の構造を比較する。原核生物では、RNAポリメラーゼによって合成されたRNAはそのままmRNAとして機能し、翻訳に使われる。したがって、mRNAの5’末端には基質となったNTP(ヌクレオシド三リン酸)の3つのリン酸基がそのまま残っている。それに対して真核生物のmRNAの5’末端には、キャップ構造として7-メチルグアノシンが付加されている。
では、その仕組みを説明しよう。
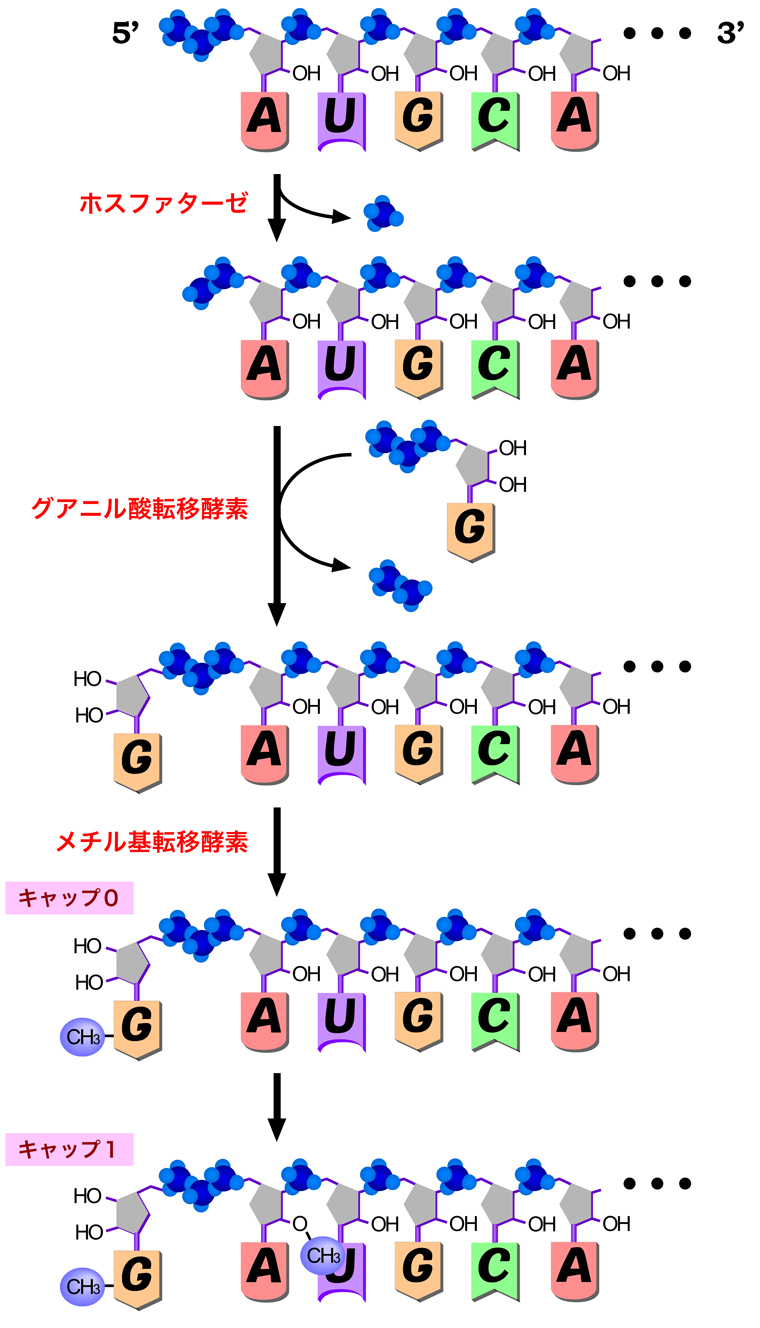 最初は原核生物と同様にRNAが合成されるため、5’末端には3つのリン酸基が残っている。そこで、RNAポリメラーゼIIによりRNAが約25ヌクレオチド合成されると、リン酸化CTDに結合していたキャッピング酵素が5’キャップ構造の形成を始める。まずホスファターゼ(脱リン酸化酵素)が作用して、末端のリン酸基を1個除去する。次にグアニル酸転移酵素が作用して、グアノシン三リン酸から二リン酸を切り取り、残ったグアノシン一リン酸をRNAの末端に付加する。通常のRNA合成では、RNAの3’末端にあるリボースの3’-OH基にヌクレオチドの5’-リン酸が付加されるが、キャップ形成のときはRNAの5’末端のリン酸基にグアノシン一リン酸のリン酸基が付加されるため、この部分ではリボースの5’と5’が三リン酸を介して向き合うことになる。そして最後に、メチル基転移酵素が付加されたグアニンの7位にメチル基を付加する。こうしてできた5’キャップ構造は『キャップ0』とよばれ、単細胞真核生物では、これが主要なキャップ構造になる。一方多細胞生物では、1番目のヌクレオチドのリボースにある2’-OHがメチル化されて『キャップ1』となったものが主になる。さらに、2番目のヌクレオチドでも2’-O-メチル化された『キャップ2』も存在する。
最初は原核生物と同様にRNAが合成されるため、5’末端には3つのリン酸基が残っている。そこで、RNAポリメラーゼIIによりRNAが約25ヌクレオチド合成されると、リン酸化CTDに結合していたキャッピング酵素が5’キャップ構造の形成を始める。まずホスファターゼ(脱リン酸化酵素)が作用して、末端のリン酸基を1個除去する。次にグアニル酸転移酵素が作用して、グアノシン三リン酸から二リン酸を切り取り、残ったグアノシン一リン酸をRNAの末端に付加する。通常のRNA合成では、RNAの3’末端にあるリボースの3’-OH基にヌクレオチドの5’-リン酸が付加されるが、キャップ形成のときはRNAの5’末端のリン酸基にグアノシン一リン酸のリン酸基が付加されるため、この部分ではリボースの5’と5’が三リン酸を介して向き合うことになる。そして最後に、メチル基転移酵素が付加されたグアニンの7位にメチル基を付加する。こうしてできた5’キャップ構造は『キャップ0』とよばれ、単細胞真核生物では、これが主要なキャップ構造になる。一方多細胞生物では、1番目のヌクレオチドのリボースにある2’-OHがメチル化されて『キャップ1』となったものが主になる。さらに、2番目のヌクレオチドでも2’-O-メチル化された『キャップ2』も存在する。
このキャップ構造は、mRNAの安定性にも影響するが、翻訳の開始でも重要な役割を果たすので、詳しくは翻訳のところで説明しよう。
RNAスプライシング
真核生物の遺伝子の約94%は、イントロンとよばれる介在配列によって分断されている。このイントロンはタンパク質をコードしない配列であるため、遺伝子のタンパク質コード領域も完全に分断されることになる。遺伝子が発現するとき、イントロンの配列はmRNA前駆体へと転写されるが、翻訳される前にはRNAスプライシングによって正確に取り除かれる。したがって、成熟mRNAにおいては、タンパク質コード領域は完全に1つにつながる。成熟mRNA中に残る配列をエキソン、成熟mRNAで切り取られる配列をイントロンという。

タンパク質をコードしていなくても、5’非翻訳領域(5’リーダー配列)や3’ 非翻訳領域は成熟mRNA中に残るため、エキソンになる。RNAスプライシングの詳細な過程は、次の頁で説明したい。
ポリA尾部の形成
原核生物の遺伝子には明確なターミネーターが存在するため、ターミネーターで転写終結したところがmRNAの3’末端となる。ところが真核生物では、mRNAの3’末端となる部分を大きく越えて転写は終結し、実際のmRNAの3’末端はRNAプロセシングにより形成される。
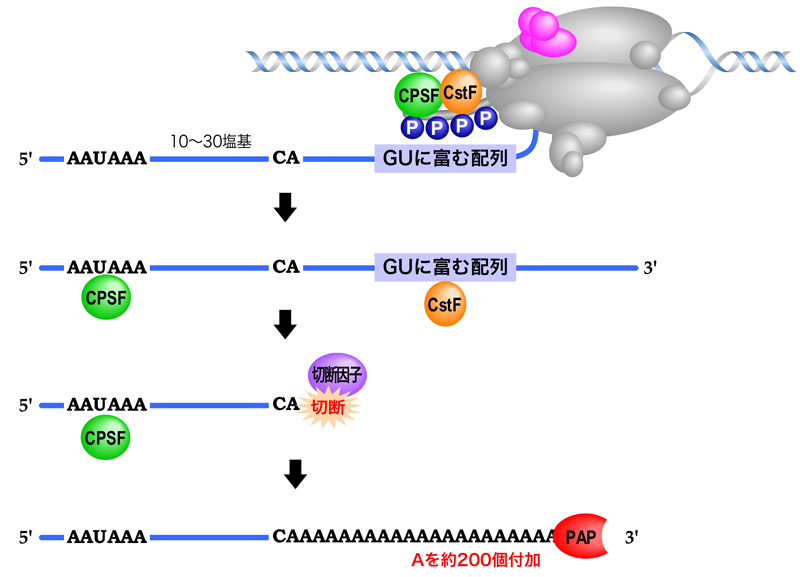
真核生物のmRNAの3’末端形成には、シグナルが存在する。これらのシグナルはゲノム中にコードされているが、RNAへと転写されたのちに機能する。まずmRNAの3’末端に近い位置にポリAシグナル(AAUAAA)があり、そこから10〜30塩基下流にCAの2塩基がある。そしてさらに下流にはGU-rich配列が存在する。
mRNA前駆体中のポリAシグナルとGU-rich配列が転写されると、リン酸化CTDに結合していたCPSF(切断ポリアデニル化特異因子)とCstF(切断促進因子F)がそれぞれ結合する。そして、切断因子がCA配列の3’側でRNA鎖を切断し、mRNA前駆体が切り離される。最後に、ポリAポリメラーゼがアデニン(A)を1個ずつ、全部で約200個付加することにより、ポリA尾部が完成する。このポリA尾部も、mRNAの安定性に重要である。
これら一連のRNAプロセシングを受けたmRNA前駆体は成熟mRNAとなる。そして、5’キャップにはキャップ結合複合体(CBC)、ポリA尾部にはポリA結合タンパク質が結合する。すべてのRNAプロセシングを適切に完了し、適切なタンパク質が結合した成熟mRNAだけが、細胞質へと輸送されて翻訳(タンパク質合成)に使われるのである。
