
このページのテーマは、これ!
どのように完全体のmRNAが作られるのか?
このページの内容は、われわれ人間を含めた真核生物にのみ当てはまります。
細菌などの原核生物には当てはまりません。
どのように完全体のmRNAが作られるのか?
このページの内容は、われわれ人間を含めた真核生物にのみ当てはまります。
細菌などの原核生物には当てはまりません。
さて、博士。今日は、転写の後の話ですね。
そうじゃ。転写が終わって、翻訳が始まるまでの話じゃよ。
転写が終わったら、mRNAが細胞質へ行って、そのまま翻訳が行われるだけではないのですか?
チッチッチ!違うんだな、これが!

ムッ!どう違うんですか?
実は、転写直後のmRNAは、完全なmRNAではないんじゃ。このページでは、この完全なmRNAが完成するまでの過程を紹介していこう。まずは、下の図を見てくれたまえ。
(注:正しくは、この完全なmRNAのことを”mRNA”と呼び、転写直後のmRNAは”hnRNA”と呼ばれる。)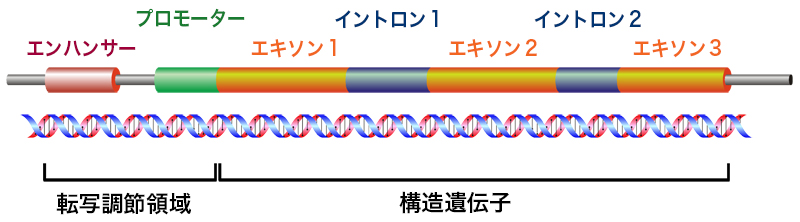
(注:正しくは、この完全なmRNAのことを”mRNA”と呼び、転写直後のmRNAは”hnRNA”と呼ばれる。)
これは、前のページでも使った図じゃ。前のページでも話したが、エクソンとは、直接的にタンパク質の構造を決定している部分じゃ。それに対して、イントロンはそれを分断している部分で、最終的には除去されてしまうんじゃ。このイントロンを除去する過程が、スプライシングと呼ばれておる。
なるほど、無駄なものは切り捨てるんですね。まさに、遺伝子界のリストラ...このスプライシングを終えて、完全なmRNAになるんですね?
いや、それだけではまだ不完全なんじゃ。スプライシング以外にも、mRNAの5’末端にキャップという構造を作ったり、3’末端にたくさんのA塩基(ポリAテール)を付加しなくてはならない。これらの一連の過程を経て、完全なmRNAが完成するんじゃ。
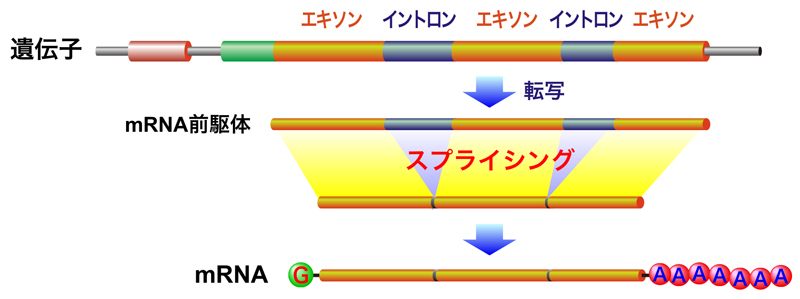
mRNAが帽子をかぶって、尻尾もつけるんですね。このページでは、完全なmRNAを作る過程について詳しく教えてくれるんですね?

うむ。まずは、スプライシングの話から行こう!スプライシングとは、上でも紹介したが、イントロンを除去してエクソンどうしをつなぎ合わせる過程じゃ。そして、イントロンの塩基配列は必ずGTで始まりAGで終わる。これをGT-AGルールというんじゃ。さらに、イントロンの中にはブランチ部位と呼ばれる塩基配列もある。
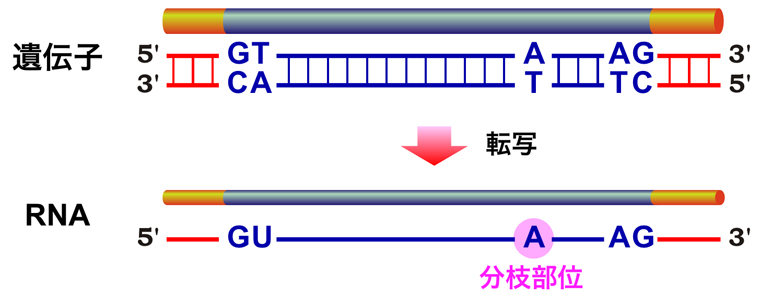
ブランチ... ここで昼食兼朝食(brunch)ですね。私はサンドイッチがいいなぁ。
違う!枝(branch)の方じゃ!分枝部位ともいう。なぜ”枝”なのかは、後で分かるじゃろう。
ふ〜ん。では、このスプライシングの時も、転写開始の時みたいに色々なタンパク質が働くんですか?
スプライシングの時に頑張るのは、主にsnRNAと呼ばれる短いRNAじゃ。このsnRNAは、普段はタンパク質と結合してsnRNPとして働く。では、snRNPたちの活躍を、順を追って説明していこう。

まず最初に、U1 snRNPがイントロンの5’末端にあるGU配列に結合する。U1 snRNP中のU1 snRNAは、mRNA中のイントロンのGU配列と塩基対を形成できるような配列を持っておるんじゃ。だから、GU配列に特異的に結合できるんじゃ。
これが、スプライシングの一番最初の段階なんですね。
続いて、U2AFというタンパク質がブランチ部位の下流に結合し、さらにU2 snRNPがブランチ部位に結合する。このU2 snRNPもU1 snRNPの場合と同様に、U2 snRNA中にブランチ部位と塩基対を形成できるような塩基配列を持っておるんじゃ。

順番にくっついていくんですね。
うむ。U1 snRNPのGU配列への結合がU2AFの結合に必要で、U1 snRNPとU2AFがU2 snRNPのブランチ部位への結合に必要なんじゃ。
なるほど、無駄がありませんねぇ。
次に、U5 snRNPとU4 snRNPとU6 snRNPも集まって、全ての因子が出そろう。そして、いよいよ本格的なスプライシングの開始じゃ。

まずは、U1 snRNPが放出される。これによって、イントロン5’末端のGU配列が開放されるんじゃ。すると、その他のU5やU6 sn RNPがこのGU配列に近づいてくる。ここで、U4 snRNPが放出されると、スプライシングは一気に活性化される。
つまり、U4 snRNPが、U6 snRNPの活性をコントロールしていたんですね?
その通りじゃ。U6 snRNPはU4 snRNPと塩基対を形成してくっついておったのじゃが、U4が放出されると、今度はU6 snRNPはU2 snRNPにくっつくんじゃよ。そして下の図のように、全体の形もダイナミックに変化するんじゃ。
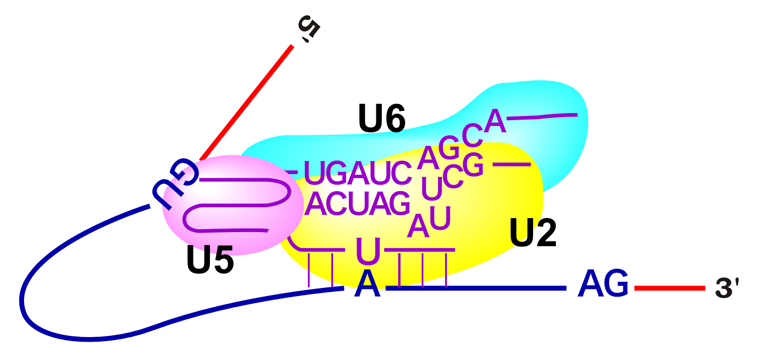
U2 snRNPも、U6 snRNPと塩基対を形成するのですか?
うむ。さらに、U6 snRNPは塩基対を利用してイントロン5’末端のGU配列とも結合するんじゃ。そして、U2/U6がGU配列の5’側でmRNAを切断し、GU配列のG塩基をブランチ部位中のA塩基と結合させるんじゃよ。
なるほど。このブランチ部位で、枝分かれしたような構造ができるから、ブランチ(枝)だったんですね。そして、カウボーイの投げ縄みたいな形になりましたね。
だから、このような構造は投げ縄構造あるいはラリアット構造と呼ばれておる。
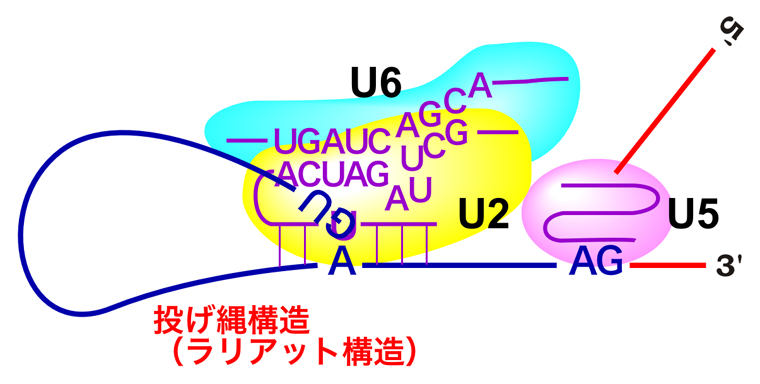
おぉ〜、スタンハンセンや長州力を彷彿とさせる名前ですねぇ。このとき、切り離されたエクソンは、フラフラしているんですか?
いや。切り離されたエクソンは、U5 snRNPがちゃんと捕まえておるんじゃ。さらにU5 snRNPは、イントロン3’末端のAG配列にも結合して、がっちり捕まえておる。だから、AG配列の3’側でmRNAが切断された後も、2つのエクソンは離れることはないんじゃよ。
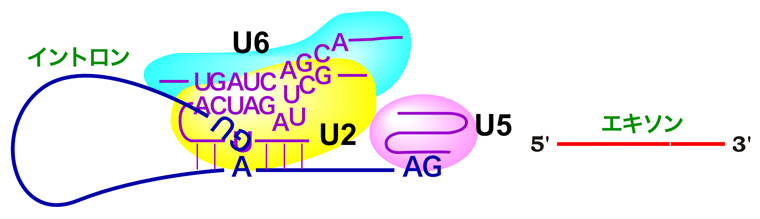
なるほど。そして最終的に2つのエクソンは繋げられるんですね。では、切り離されたイントロンはどうなるんですか?
このイントロンは、投げ縄構造を形成したまま放出されるが、すぐにまたまっすぐな1本の鎖に戻り、すぐに分解されてしまうんじゃ。そして、以上のスプライシングの過程をまとめると、下のアニメーションのようになる。

ふ〜ん。うまくできているんですね。ところで、最初に出てきたキャップとポリAテールは、何をしているんですか?
うむ。一般的に、キャップ構造はmRNAが5’末端から分解されるのを保護し、翻訳の開始に重要な役割を持っていると言われておる。 そしてポリAテールは、mRNAの安定性と関連があると言われておるんじゃ。
なるほど。これで、完全体のmRNAの完成なんですね。
その通りじゃ。何か他に質問はあるかね?
はい。ウェスタンラリアットと力ラリアットの違いは何ですか?
...
